Výber čitateľov

Populárne články
V ochranných sietí a mnoho zvierat obsahovalo pigment rhodopsinalebo vizuálne purpur, zloženie, vlastnosti a chemické transformácie, ktoré boli podrobne študované v posledných desaťročiach. Pigment bol nájdený v Kolodsku jódpcín. V Kolodnok sú tu tiež pigmenty chlorolabu a erytrolabu; Prvá z nich absorbuje lúče zodpovedajúce zelenej a druhej - červenej časti spektra.
Rhodopsin Je to vysoko molekulárna zlúčenina (molekulová hmotnosť 270 000) pozostávajúca zo sietnice - aldehyd vitamínu A a lúča opciny. Pod pôsobením fúzie Quantum sa vyskytuje cyklus fotofyzikálnych a fotochemických transformácií tejto látky: retinal je úžasný, jeho bočný reťazec je znázornený, spojenie sietnika s proteínom je rozbité, aktivujú sa enzymatické centrá proteínovej molekuly. Potom sa retinálny retinál štiepi z opcínu. Pod vplyvom enzýmu nazývaného reduktázy retinal, druhý sa pohybuje do vitamínu A.
Keď sa oko stmieva, regenerácia vizuálnej fialovej sa vyskytuje, t.j. Resintez Rhodopsin. Pre tento proces je potrebné, aby sietnica dostala cis-izomér vitamínu A, z ktorej sa vytvorí sietnika. Ak neexistuje vitamín A v tele, tvorba Rhodopsínu je prudko porušená, čo vedie k rozvoju kine slepoty.
Fotochemické procesy v sietnici sa vyskytujú veľmi ekonomicky, t.j. Pod krokom, dokonca aj veľmi jasné svetlo rozdelí len malú časť rhodopcínových prútikov.
Štruktúra yodopcínu je blízko Rhodopsínu. Jodopcín je tiež zmesi spojenia s proteínom, ktorý je vytvorený v stĺpcoch a líši sa z pečiatok tyčiniek.
Absorpcia ľahkého rhodopsínu a jóxínu je odlišná. Jodopcín je najviac absorbovaný žltým svetlom s vlnovou dĺžkou približne 560 nm.
Na dlhej vlnovej hranici viditeľného spektra sú lúče červenej (vlnovej dĺžky 723-647 nm), na krátkom čase - fialová (vlnová dĺžka 424-397 nm). Miešanie lúčov všetkých spektrálnych farieb dáva bielu farbu. Biela farba je možné získať a pri zmiešaní dvoch takzvaných párových voliteľných farieb: červená a modrá, žltá a modrá. Ak miešate farby z rôznych párov, môžete získať medziľahlé farby. V dôsledku zmiešania troch hlavných farieb spektra - červenej, zelenej a modrej - môžu byť získané akékoľvek farby.
Farbenie teórií. Existuje množstvo teórií farieb, trojzložková teória využíva najväčšie uznanie. Tvrdí, že existuje v sietnici troch rôznych vrchov farebných fotoreceptorov - Kollok.
Na existenciu trojzložkového mechanizmu vnímania kvetov, povedal M. V. Lomonosov. V budúcnosti bola táto teória formulovaná v roku 1801 T. Jung a G. Helmholz je vyvinutý. Podľa tejto teórie sú stĺpce rôzne fotosenzitívne látky. Niektoré stĺpce obsahujú látku, ktorá je citlivá na červenú, iné - zelené, tretie - do fialovej. Každá farba má akciu na všetkých troch farebných prvkach, ale v rôznych stupňoch. Tieto excitations sú zhrnuté s vizuálnymi neurónmi a dosahujú kôru, dávajú pocit jednej alebo inej farby.
Podľa inej teórie navrhnutej E. Gersom sú tri hypotetické fotosenzitívne látky v stĺpcoch mesh: biela-čierna, červená zelená a žltá-modrá. Rozpad týchto látok pod vplyvom svetla vedie k pocitu bielej, červenej alebo žltej. Ostatné ľahké lúče spôsobujú syntézu týchto hypotetických látok, v dôsledku čoho sa objaví pocit čiernej, zelenej a modrej.
Najvhodnejšie potvrdenie v elektrofyziologických štúdiách získalo trojzložkovú teóriu farieb. V experimentoch na zvieratách s použitím mikroelektródami boli impulzy pridelené z jediných buniek sietnice ganglion, keď osvetľujú rôznymi monochromatickými lúčmi. Ukázalo sa, že elektrická aktivita vo väčšine neurónov sa vyskytla pod pôsobením lúčov akejkoľvek vlnovej dĺžky vo viditeľnej časti spektra. Takéto retinálne prvky sú menované Dominators. V ostatných gangliových bunkách (modulátoroch) sa impulzy vyskytujú len pri osvetlení lúčov len určitú vlnovú dĺžku. 7 modulátorov bolo zistených, že optimálne reagujú na svetlo s rôznymi vlnovými dĺžkami (od 400 do 600 nm). R. GRANITE sa domnieva, že tri zložky farieb vnímania, ktoré boli suspendované T. Jungom a Gelmholtzom, sa získajú spriemerovaním kriviek spektrálnej citlivosti modulátorov, ktoré môžu byť zoskupené podľa troch hlavných častí spektra: modrá fialová , zelená a oranžová.
Pri meraní microfectreerometrometra absorpcie lúčov rôznych vlnových dĺžok s jedným clummerom sa ukázalo, že niektoré stĺpce absorbujú červené oranžové žiarenie čo najviac, druhá - Tretia je modré lúče. Tri skupiny plodov sú teda odhalené v sietnici, z ktorých každý vníma lúče zodpovedajúce jednému z hlavných farieb spektra.
Trojzložková teória farebnej vízie vysvetľuje množstvo psycho-fyziologických javov, ako je napríklad postupné farebné obrazy a niektoré fakty farebnej patológie (slepota s ohľadom na jednotlivé farby). V posledných rokoch sa v sietnici a vizuálnych centrách skúmali mnoho tzv. Oponentov neurónov. Líšia sa tým, že účinok na oko žiarenia v určitej časti spektra je vzrušený a v iných častiach spektra - spomaľuje. Predpokladá sa, že takéto neuróny najúčinnejšie kódujú informácie o farbe.
Farbosleposť. Daltonizmus sa vyskytuje u 8% mužov, výskyt je spôsobený genetickým nedostatkom určitých génov v rozhodujúcom podlahe nepárového X-chromozómu. Aby sa diagnostikovať daltonizmus, test naznačuje sériu polychromatických tabuliek alebo zadávať farebné jednotlivé objekty rôznych farieb. Diagnóza Daltonizmu je dôležitá v profesionálnom výbere. Ľudia trpiacim Daltonizmom nemôžu byť vodičmi dopravy, pretože nerozlišujú farby semaforov.
Existujú tri odrody čiastkovej farebnej suspenzie: protoropy, Deuteropia a Tritonia. Každý z nich je charakterizovaný neprítomnosťou vnímania jednej z troch hlavných farieb. Ľudia trpiaci protoropia ("Kitty") nie sú vnímané červeno, modro-modré lúče sa zdajú byť bezfarebné. Osoby, ktoré trpia determansopy ("zelená-zastávka) nerozlišujú zelené farby z tmavej červenej a modrej. S Tritonia - zriedka sa stretol s anomáliami farebného videnia, lúče modrej a fialovej nie sú vnímané.
Vizuálny analyzátor je kombináciou štruktúr, ktoré vnímajú svetelnú energiu vo forme elektromagnetického žiarenia s vlnovou dĺžkou 400 - 700 nm a diskrétnymi časticami fotónov alebo kvanta a tvoriace vizuálne pocity. S pomocou oka vnímaná 80-90% všetkých informácií o celom svete.
Vďaka aktivite vizuálneho analyzátora sa rozlišuje osvetlenie objektov, ich farba, tvar, veľkosť, smer pohybu, vzdialenosť, ku ktorej sú odstránené z oka a od seba. To všetko vám umožní odhadnúť priestor, orient na svete po celom svete, vykonávať rôzne typy zameraných aktivít.
Spolu s koncepciou vizuálneho analyzátora je koncept zorného orgánu.
Orgán z pohľadu je oko, ktoré obsahuje tri rôzne vo funkčnosti prvku:
Ø Eye Eye Apple, v ktorom sú umiestnené svetelné skrine a zariadenia na kontrolu svetla;
Ø ochranné zariadenia, t.j. vonkajšie plášte (sklera a rohovka), trhlové prístroje, očné viečka, riasy, obočie;
Ø Musculosor prezentovaný tromi výpary očných svalov (vonkajšie a vnútorné priame, hornej a dolnej rovnej, hornej a dolnej časti šikmé), ktoré sú inervované III (otvárací nerv), IV (blokový nerv) a VI (čerpací nerv) s lebečným nervom párov.
Štrukturálne a funkčné charakteristiky
Oddelenie receptora (periférne) oddelenie vizuálneho analyzátora (fotoreceptory) je rozdelené na laná a columín neurosenzorské bunky, ktorých vonkajšie segmenty, ktoré sú valcované ("palice") a columine ("copumy") formy. Osoba má 6-7 miliónov stĺpcov a 110 - 125 miliónov oteckov.
Miesto konania vizuálneho nervu z sietnice neobsahuje fotoreceptory a nazýva sa slepý bod. Bočné z slepého miesta v strede centrálneho piateho leží miesto na najlepšie vízie - žlté miesto obsahujúce prevažne Kolkochki. Na okraj sietnice sa počet plutiev znižuje a počet sa zvyšuje počet tyčiniek a periféria sietnice obsahuje iba prútiky.
Rozdiely vo funkciách kolónov a tyčiniek sú základom fenoménu duality videnie. Tyčinky sú receptory, ktoré vnímajú svetelné lúče v podmienkach nízkeho svetla, to znamená bezfarebné, alebo achromatické, vízie. Stĺpce fungujú vo svetlom osvetlení a sú charakterizované rôznou citlivosťou na spektrálne vlastnosti svetla (farebné alebo chromatické videnie). Fotoreceptory majú veľmi vysokú citlivosť, vďaka znaku štruktúry receptorov a fyzikálno-chemických procesov, ktoré sú základom vnímania energie svetelného stimulu. Predpokladá sa, že fotoreceptory sú nadšené pod akciou na nich 1 - 2 svetelný kvantón.
Tyčinky a stĺpce pozostávajú z dvoch segmentov - vonkajšie a vnútorné, ktoré sú spojené pomocou úzkych cilia. Tyčinky a stĺpce sú orientované v sietnici radiálne a molekuly fotosenzitívnych proteínov sú umiestnené vo vonkajších segmentoch takým spôsobom, že asi 90% ich fotosenzitívnych skupín leží v rovine diskov obsiahnutých v vonkajších segmentoch. Svetlo má najväčší vzrušujúci účinok, ak sme smer lúča zhoduje s dlhou osou prútika alebo misy, zatiaľ čo je nasmerovaná kolmo na disky svojich vonkajších segmentov.
Fotochemické procesy v očiach sietnice. V receptorových bunkách sietnice sú fotosenzitívne pigmenty (komplexné proteínové látky) - chromoproteidy, ktoré sú sfarbené vo svetle. V paličkach na membráne vonkajších segmentov, Rhodopsin je obsiahnutý v Kolodskoku - jódpcín a iných pigmentoch.
Rhodopcín a jódpcín pozostávajú zo sietnice (aldehyd vitamín A1) a glykoproteín (OCTIC). Mať podobnosť v fotochemických procesoch, sa líšia tým, že maximálna absorpcia je v rôznych oblastiach spektra. Choptiny obsahujúce Rhodopsin majú maximálnu absorpciu v oblasti 500 nm. Existujú tri typy medzi plemami, ktoré sa líšia v maximá v absorpčnom spektre: niektoré majú maximum modrej časti spektra (430 - 470 nm), iní v zelenej (500 - 530), tretí - červená (620 - 760 nm) časti, ktoré sú spôsobené prítomnosťou troch typov vizuálnych pigmentov. Červený pigment Colummer sa nazýva "jódpcín". Retinál môže byť v rôznych priestorových konfiguráciách (izomérnych foriem), ale iba jeden z nich je 11-cis-izomér retinalu, pôsobí ako chromofórová skupina všetkých známych vizuálnych pigmentov. Zdrojom sietnice v tele je karotenoidy.
Fotochemické procesy v sietnici pokračujú veľmi ekonomicky. Dokonca aj pri pôsobení jasného svetla sa štiepi len malá časť rhodopcínu existujúceho v paličkach (asi 0,006%).
V tme sa obnoví pigmenty prúdiace s absorpciou energie. Reštaurovanie jodopcínu prúdi 530 krát rýchlejšie ako Rhodopsina. Ak organizmus znižuje obsah vitamínu A, procesy opätovného zavedenia Rhodopsínu oslabujú, čo vedie k porušeniu videnie Twilight, tzv. Kuracie slepoty. S konštantným a rovnomerným osvetlením je rovnováha medzi rýchlosťou rozpadu a opätovným zavedením pigmentov. Keď sa množstvo svetla pádu na sietnici zníži, táto dynamická rovnováha je rozbitá a posunutá smerom k vyšším koncentráciám pigmentov. Tento fotochemický fenomén je základom temnej adaptácie.
Osobitne dôležitý význam vo fotochemických procesoch má retinálnu pigmentovú vrstvu, ktorá je tvorená epitelom obsahujúcim fuscine. Tento pigment absorbuje svetlo, prevenciu reflexie a disperzie, čo spôsobuje jasnosť vizuálneho vnímania. Pigmentové bunky procesy obklopujú fotosenzitívne segmenty palice a kolódy, ktoré sa zúčastňujú na výmene látok fotoreceptorov a syntézy vizuálnych pigmentov.
Vzhľadom na fotochemické procesy vo fotoreceptoroch oka v pôsobení svetla, dochádza k receptorovému potenciálu, čo je hyperpolarizácia receptorovej membrány. Ide o výrazný znak vizuálnych receptorov, aktivácia iných receptorov je exprimovaná vo forme depolarizácie ich membrány. Amplitúda potenciálu vizuálneho receptora sa zvyšuje s zvyšovaním intenzity svetelného stimulu. Takže v pôsobení červenej, vlnovej dĺžky, ktorej je 620-760 nm, receptorový potenciál je výraznejší v fotoreceptoroch centrálnej časti sietnice a modrá (430 - 470 nm) je v periférii.
Synaptické zakončenie fotoreceptora konvep na bipolárnych sietnických neurónoch. Súčasne sú fotoreceptory centrálnych jamiek spojené len s jedným bipolárnym. Vedúce oddelenie vizuálneho analyzátora začína z bipolárnych buniek, potom ganglion buniek, potom vizuálny nerv, potom vizuálne informácie vstupuje do bočných kľukových hriadeľov thalamu, odkiaľ sa v zložení vizuálneho žiarenia premieta do primárnych vizuálnych polí.
Primárnym vizuálnym poliam Cortex je pole 16 a pole 17 - to je špongia drážka o týlných akciách. Osoba je charakterizovaná binokulárnymi stereoskopickým víziou, to znamená, že schopnosť rozlíšiť objem objektu a zobrazený dve oči. Adaptácia svetla je charakteristická, to znamená zariadenie pre určité svetelné podmienky.
Po dlhú dobu je známy fenomén luminiscencie - látka absorbuje svetlo určitej frekvencie a samotná vytvára rozptýlený N (lepšia frekvencia. Stále v XIX storočí. Stokes Nastavte pravidlo - frekvencia rozptýleného svetla je menej ako frekvencia absorbovaného (ν pravidiel\u003e ν rady); fenomén sa vyskytuje len vtedy, keď často vysoká frekvencia padajúceho svetla.
V niektorých prípadoch sa Luminincescence vyskytuje takmer najjemnejší - okamžite sa vyskytuje a zastavuje sa po 10 -7 -10 -8 s po ukončení osvetlenia. Tento konkrétny prípad luminiscencie sa niekedy volá fluorescencia. Mnohé látky (fosforu a iné) však majú dlhý rozkrok, trvalý (postupne oslabujúci) minúty a dokonca aj hodiny. Tento druh luminiscencie bol volaný fosforescencie. Po zohriatí sa telo stráca schopnosť fosforečinu, ale zachováva schopnosť lumínu.
Vynásobenie oboch častí nerovnosti vyjadrujúcej pravidlo Stokes, na stálej doske, dostaneme:
V dôsledku toho fotónová energia absorbovaná atómom, viac energie emitovaného fotónu; A tu sa prejavuje fotónová povaha procesov absorpcie svetla.
Dospeli sme dostupné odchýlky od pravidla Stokes neskôr (§ 10.6).
Vo fenoménoch fotochémie - chemické reakcie v pôsobení svetla, bolo tiež možné stanoviť existenciu najmenšej frekvencie, ktorá sa má vyskytnúť. To je dosť zrozumiteľné z pohľadu fotónov: pre výskyt reakcie molekuly by mal získať dostatočnú ďalšiu energiu. Často je fenomén maskovaný ďalšími účinkami. Je známe, že zmes vodíka H2 s chlórom Cl2 v tme je dlhý čas. Ale aj so slabým osvetlením s ľahkou vysokou frekvenciou, zmes veľmi rýchlo exploduje.
Dôvod spočíva v výskyte sekundárnych reakcií. Molekula vodíka, absorbuje fotón, môže disociovať (hlavnú reakciu):
H 2 + Hν -\u003e N + N.
Pretože atómový vodík je oveľa aktívnejší ako molekulárny, potom sekundárna reakcia vzniká s uvoľňovaním tepla:
H + SL 2 \u003d HCL + SL.
Atómy N a Cl sú teda vyňaté. Interagujú s molekulami C12 a H2 a reakcia rastie veľmi násilne, je jeden deň nadšený absorpciou malého počtu fotónov.

Medzi rôzne fotochemické reakcie, reakcie, ktoré sa konajú, sú zaslúžené na procese fotografií. Fotoaparát vytvára platný (zvyčajne redukovaný) obrazu na fotografickej emulznej vrstve obsahujúcej strieborný bromid schopný fotochemických reakcií. Počet molekúl odpovedí približne približne úmernej intenzity svetla a času jej pôsobenia (dĺžka expozície pri fotografovaní). Toto číslo je však relatívne veľmi malé; Výsledný "skrytý obraz" sa podrobí procesu prejavu, keď pod pôsobením zodpovedajúcich chemických činidiel existuje ďalší selekcia bromidu striebra v centrách pochádzajúcich počas fotochemickej reakcie. Potom nasleduje proces upevnenia (upevnenie) obrazu: nezreagovaný silver bromid je preložený do roztoku a kovové striebro zostáva na fotoshlení, čo určuje priehľadnosť jednotlivých častí získaného negatívneho obrazu (čím viac sa svetlo absorbuje, tmavšie zodpovedajúceho miesta). Osvetlenie fotografického papiera (alebo filmu) cez negatívne, sa dostanú na papier (po jeho prejave a fixácii) distribúcia osvetlenia zodpovedajúce odnímateľnému objektu (samozrejme, ak sú splnené vhodné podmienky na snímanie a spracovanie fotografického materiálu) . Keď farebná fotografia, film obsahuje tri vrstvy citlivé na tri rôzne časti spektra.
Tieto vrstvy slúžia ako ľahké filtre pre seba a osvetlenie každého z nich je určené len špecifickou časťou spektra. Byť oveľa zložitejším ako čiernobiely foto proces, farebný proces fotografovania v zásade sa nelíši od prvého a je typický fotonický proces.
Vizuálny analyzátor je kombináciou štruktúr, ktoré vnímajú svetelnú energiu vo forme elektromagnetického žiarenia s vlnovou dĺžkou 400-700 nm a diskrétnymi časticami fotónov alebo kvanta a tvoria vizuálne pocity. S pomocou oka vnímaná 80 - 90% všetkých informácií o svete okolo.
Obr. 2.1
Vďaka aktivite vizuálneho analyzátora sa rozlišuje osvetlenie objektov, ich farba, tvar, veľkosť, smer pohybu, vzdialenosť, ku ktorej sú odstránené z oka a od seba. To všetko vám umožní odhadnúť priestor, orient na svete po celom svete, vykonávať rôzne typy zameraných aktivít.
Spolu s konceptom vizuálneho analyzátora je koncept zorného orgánu (obr. 2.1)
Toto je oko, ktoré obsahuje tri rôzne prvky v funkčnosti:
1) očná guľa, v ktorej sú umiestnené svetelné priechody, svetlo načasovanie a svetelné zariadenia;
2) Ochranné zariadenia, t.j. vonkajšie plášte oka (sklerra a rohovka), trhlové prístroje, viečka, riasy, obočie; 3) Motorové zariadenie reprezentované tromi výpary oka svalov (vonkajšie a vnútorné priame, hornej a dolnej rovnej, hornej a dolnej šikmej), ktoré sú inervované III (ooo oko), IV (blokový nerv) a VI (nerv vozidla) s kraniálnymi nervovými pármi.
Štrukturálne a funkčné charakteristiky
Oddelenie receptora (periférneho) vizuálny analyzátor (fotoreceptory) je rozdelený na laná a columín neurosenzorové bunky, ktorých vonkajšie segmenty sú valcovanie ("tyčinky") a colummoid ("stĺpce") formy. Osoba má 6 - 7 miliónov stĺpcov a gule 110-125 miliónov prúžok.
Miesto konania vizuálneho nervu z sietnice neobsahuje fotoreceptory a nazýva sa slepý bod. Bočné z slepého miesta v strede centrálneho piateho leží miesto na najlepšie vízie - žlté miesto obsahujúce prevažne Kolkochki. Na okraj sietnice sa počet plutiev znižuje a počet sa zvyšuje počet tyčiniek a periféria sietnice obsahuje iba prútiky.
Rozdiely vo funkciách kolónov a tyčiniek sú základom fenoménu duality videnie. Tyčinky sú receptory, ktoré vnímajú svetelné lúče v podmienkach nízkeho svetla, t.j. Bezfarebné alebo achromatické, vízie. Stĺpce fungujú vo svetlom osvetlení a sú charakterizované rôznou citlivosťou na spektrálne vlastnosti svetla (farebné alebo chromatické videnie). Fotoreceptory majú veľmi vysokú citlivosť, vďaka znaku štruktúry receptorov a fyzikálno-chemických procesov, ktoré sú základom vnímania energie svetelného stimulu. Predpokladá sa, že fotoreceptory sú nadšené pod akciou na nich 1-2 Quanta Light.
Tyčinky a stĺpce pozostávajú z dvoch segmentov - vonkajšie a vnútorné, ktoré sú spojené pomocou úzkych cilia. Tyčinky a stĺpce sú orientované v sietnici radiálne a molekuly fotosenzitívnych proteínov sú umiestnené vo vonkajších segmentoch takým spôsobom, že asi 90% ich fotosenzitívnych skupín leží v rovine diskov obsiahnutých v vonkajších segmentoch. Svetlo má najväčší vzrušujúci účinok, ak sme smer lúča zhoduje s dlhou osou prútika alebo misy, zatiaľ čo je nasmerovaná kolmo na disky svojich vonkajších segmentov.
Fotochemické procesy v očiach sietnice.V receptorových bunkách sietnice sú fotosenzitívne pigmenty (komplexné proteínové látky) - chromoproteidy, ktoré sú sfarbené vo svetle. V paličkach na membráne vonkajších segmentov, Rhodopsin je obsiahnutý v Kolodskoku - jódpcín a iných pigmentoch.
Rhodopsin a jódpcín pozostávajú zo sietnice (aldehyd vitamín A 1) a glykoproteín (OCTIC). Mať podobnosť v fotochemických procesoch, sa líšia tým, že maximálna absorpcia je v rôznych oblastiach spektra. Choptiny obsahujúce Rhodopsin majú maximálnu absorpciu v oblasti 500 nm. Existujú tri typy medzi sprievodcami, ktoré sa vyznačujú Maxima v absorpčnom spektre: niektoré majú maximum modrej časti spektra (430-470 nm), iní v zelenej (500-530), tretí - v červenej farbe (620-760 nm) časti z dôvodu prítomnosti troch typov vizuálnych pigmentov. Červený pigment Colummer sa nazýva "jódpcín". Retinál môže byť v rôznych priestorových konfiguráciách (izomérnych foriem), ale iba jeden z nich je 11-cis-izomér retinalu, pôsobí ako chromofórová skupina všetkých známych vizuálnych pigmentov. Zdrojom sietnice v tele je karotenoidy.
Fotochemické procesy v sietnici pokračujú veľmi ekonomicky. Dokonca aj pri pôsobení jasného svetla sa štiepi len malá časť rhodopcínu existujúceho v paličkach (približne 0,006%).
V tme sa obnoví pigmenty prúdiace s absorpciou energie. Reštaurovanie jodopcínu prúdi 530 krát rýchlejšie ako Rhodopsina. Ak organizmus znižuje obsah vitamínu A, procesy opätovného zavedenia Rhodopsínu oslabujú, čo vedie k porušeniu videnie Twilight, tzv. Kuracie slepoty. S konštantným a rovnomerným osvetlením je rovnováha medzi rýchlosťou rozpadu a opätovným zavedením pigmentov. Keď sa množstvo svetla pádu na sietnici zníži, táto dynamická rovnováha je rozbitá a posunutá smerom k vyšším koncentráciám pigmentov. Tento fotochemický fenomén je základom temnej adaptácie.
Osobitne dôležitý význam vo fotochemických procesoch má retinálnu pigmentovú vrstvu, ktorá je tvorená epitelom obsahujúcim fuscine. Tento pigment absorbuje svetlo, prevenciu reflexie a disperzie, čo spôsobuje jasnosť vizuálneho vnímania. Pigmentové bunky procesy obklopujú fotosenzitívne segmenty palice a kolódy, ktoré sa zúčastňujú na výmene látok fotoreceptorov a syntézy vizuálnych pigmentov.
Vzhľadom na fotochemické procesy vo fotoreceptoroch oka v pôsobení svetla, dochádza k receptorovému potenciálu, čo je hyperpolarizácia receptorovej membrány. Ide o výrazný znak vizuálnych receptorov, aktivácia iných receptorov je exprimovaná vo forme depolarizácie ich membrány. Amplitúda potenciálu vizuálneho receptora sa zvyšuje s zvyšovaním intenzity svetelného stimulu. Takže v pôsobení červenej, vlnovej dĺžky, ktorej je 620-760 nm, receptorový potenciál je výraznejší v fotoreceptoroch centrálnej časti sietnice a modrá (430-470 nm) je v perifériách.
Synaptické zakončenie fotoreceptora konvep na bipolárnych sietnických neurónoch. Súčasne sú fotoreceptory centrálnych jamiek spojené len s jedným bipolárnym.
Vodivé oddelenie.Prvým neurónom vedenia oddelenia vizuálneho analyzátora je reprezentovaný bipolárnymi retinálnymi bunkami (obr. 2.2).

Obr. 2.2.
Predpokladá sa, že v bipolárnych bunkách existujú potenciály účinku podobné receptorom a horizontálni. V niektorých bipolárnych sa pomaly dlhodobá depolarizácia vyskytuje v niektorých bipolárnych zaniknutých a vypnutých a v iných - na inklúziu - hyperpolarizácia, pri odstavení - depolarizácia.
Axóny bipolárnych buniek sa zase konvertujú na gangliové bunky (druhý neurón). Výsledkom je, že približne 140 tyčiniek sa môže konvergovať pre každú ganglionovú bunku, zatiaľ čo bližšie k žltého miesta, tým menšie fotoreceptory spoznávajú rovnakú bunku. V oblasti žltých škvŕn sa konvergencia takmer nevykonáva a počet plodov sa takmer rovná počtu bipolárnych a gangliových buniek. To je presne, ako vysoká zraková ostrosť vysvetľuje v centrálnych retinálnych oddeleniach.
Okrese sietnice sa vyznačuje veľkou citlivosťou na slabé svetlo. To je pravdepodobne spôsobené tým, že až 600 tyčiniek sa tu konvertuje cez bipolárne bunky na rovnakej gangliovej bunke. Výsledkom je, že signály zo súboru palice sú zhrnuté a spôsobujú intenzívnejšiu stimuláciu týchto buniek.
V gangliových bunkách sa vygeneruje séria impulzov s frekvenciou 5 za sekundu. Táto impulzia sa deteguje s mikroelektródou štúdie jednotlivých vizuálnych vlákien alebo jednotlivých gangliových buniek a v tme je vnímané ako "vlastné očné svetlo".
V niektorých bunkách ganglion dochádza na účasť výtlačku pozadia na svetle na (on-odpoveď), v iných - vypnúť svetlo (off-odpoveď). Reakcia ganglion bunky môže byť spôsobená spektrálnym zložením svetla.
V sietnici, okrem zvisle, existujú aj bočné spojenia. Bočné interakcie receptorov sa uskutočňuje horizontálnymi bunkami. Bipolárne a ganglidové bunky navzájom vzájomne pôsobia vďaka mnohým bočným väzbám tvoreným kolaterálom dendritov a axónov samotných buniek, ako aj pomocou buniek amacrínových buniek.
Horizontálne retinálne bunky poskytujú reguláciu prenosu pulzov medzi fotoreceptormi a bipolárnym, reguláciou vnímania farieb a prispôsobením oka na rôzne osvetlenie. Počas celého osvetľovacieho obdobia horizontálne bunky vytvárajú pozitívny potenciál - pomalú hyperpolarizáciu, nazývanú S-potenciál (z angličtiny - pomaly). Pri povahe vnímania podráždenia svetla sú horizontálne bunky rozdelené do dvoch typov:
1) typ L, v ktorom sa S-potenciál vyskytuje pod pôsobením akejkoľvek vlny viditeľného svetla;
2) Typ C alebo "Farba", typ, v ktorom sa potenciálne odchýlkové znamenie závisí od vlnovej dĺžky. Takže červené svetlo môže spôsobiť ich depolarizáciu a modrá je hyperpolarizácia.
Predpokladá sa, že horizontálne bunkové signály sa prenášajú v elektroteronskej forme.
Horizontálne, ako aj amecrínové bunky sa nazývajú brzdové neuróny, pretože poskytujú bočné brzdenie medzi bipolárnymi alebo gangálnymi bunkami.
Kombinácia fotografických sedadiel posielajúcich svoje signály na jeden ganglion bunky tvorí svoje pole recept. V blízkosti žltej škvrny majú tieto polia priemer 7-200 nm, a na periférii - 400-700 nm, t.j. V strede sietnice sú vnímavé polia malé, a na obvode sietnice sú oveľa väčšie ako priemer. Retinálne receptívne polia majú zaoblený tvar, koncentricky konštruovaný, každý z nich má excitačné centrum a brzdnú periférnu zónu vo forme kruhu. Existujú vnímavé polia s centrom (vzrušený, keď stredové osvetlenie) a z kancelárie (nadšený počas stmavnutia stredu). Brzdové krivky, ako naznačujú v súčasnosti, tvoria horizontálne články sietnice na mechanizme bočného brzdenia, t.j. Čím silnejší je stred receptívneho poľa vzrušený, väčší brzdný účinok má perifériu. Vďaka takýmto typom polí receptov (RP) gangliových buniek (s on-a off-centrami), svetlo a tmavé objekty sú detekované v oblasti pohľadu na úrovni sietnice.
V prítomnosti zvierat sa rozlišuje farba farby RP ganglionických buniek sietnice. Táto organizácia je, že určitá gangliová bunka prijíma vzrušujúce a brzdové signály z Kolkus, ktoré majú rôznu spektrálnu citlivosť. Ak napríklad "červené" stĺpce majú vzrušujúci účinok na túto gangliovú bunku, potom sa spomaľujú "modré" stĺpce. Zistili sa rôzne kombinácie vzrušujúcich a brzdných vstupov z rôznych tried. Významná časť farebných popponentných buniek ganglion sú spojené so všetkými tromi typmi hrubého čreva. V dôsledku takejto organizácie RP sa jednotlivé gangliové bunky stávajú voličom na osvetlenie určitej spektrálnej kompozície. Takže, ak vzrušenie vzniká z "červených" colums, potom excitácia modrých a zeleno-dobre citlivých pleľôčok spôsobí brzdenie týchto buniek, a ak je ganglion bunka nadšená z čarodejníkov s modromi, potom je to inhibované zeleným a červeným citlivým, atď.

Obr. 2.3.
Centrum a periféria receptívneho poľa majú maximálnu citlivosť v opačných koncoch spektra. Takže, ak centrum receptívneho poľa zodpovedá zmene aktivity na zahrnutie červeného svetla, periféria reaguje na zahrnutie modrej. Mnohé ganglion retinálne ganglidové bunky má takzvanú smerovú citlivosť. To sa prejavuje v tom, že keď sa stimul pohybuje v jednom smere (optimálny), gangliová bunka je aktivovaná, s iným smerom pohybu - neexistuje žiadna reakcia. Predpokladá sa, že selektivita reakcií týchto buniek do pohybu v rôznych smeroch je vytvorená horizontálnymi bunkami, ktoré majú podlhovasté spôsoby (tele-dendritov), \u200b\u200bs pomocou ktorých uniknú gangliové bunky. Kvôli konvergencii a bočným interakciám sa prekryjú poliam susedných gangliových buniek. To spôsobuje možnosť súhrnu účinkov ľahkých vplyvov a výskytu vzájomných vzťahov brzdy v sietnici.
Elektrické javy v sietnici. V sietnici oka, kde je oddelenie receptora vizuálneho analyzátora lokalizované a oddelenie vedenia začína, v reakcii na akciu svetla, sa vyskytujú komplexné elektrochemické procesy, ktoré môžu byť registrované ako celková odpoveď - elektrické sinogramy (ERG) (Obr. 2.3).
ERG odráža vlastnosti svetelného stimulu ako farby, intenzitu a trvanie jeho účinku. ERG môže byť registrovaná z celého oka alebo priamo z sietnice. Ak chcete získať, je jedna elektróda umiestnená na povrchu nadržanej škrupiny a druhý sa aplikuje na kožu tváre v blízkosti oka alebo na Uhmoch.
V ERG, registrovaných pri osvetľovaní oka, niekoľko charakteristických vĺn sa líši. Prvá negatívna vlna A je malá amplitúda elektrickej oscilácie, ktorá odráža excitácia fotoreceptorov a horizontálnych buniek. Rýchlo sa pohybuje na strmú-crack pozitívnu vlnu B, ktorá sa vyskytuje v dôsledku excitácie bipolárnych a amacrínových buniek. Po vlny B je pomalou elektropozitívnou vlnou C výsledkom excitácie pigmentových epitelových buniek. S okamihom ukončenia podráždenia svetla, vzhľad elektropozitívnej vlny D je viazanie.
Ukazovatele ERG sú široko používané na klinike očných ochorení na diagnostiku a kontrolu liečby rôznych ochorení oka spojeného s poškodením sietnice.
Vodivé oddelenie začínajúce v sietnici (prvý neurón je bipolárny, druhý neurónový ganglion bunky), anatomicky sa ďalej prezentuje vizuálnymi nervmi a po čiastočnom prechode ich vlákien - vizuálne trakty. V každom vizuálnom trakte sú nervové vlákna obsiahnuté z vnútorného (nosného) povrchu sietnice na tej istej strane a z vonkajšej polovice sietnice iného oka. Vlákna vizuálneho traktu sa posielajú do vizuálnej Bugarh (vlastne talamus), metatalamus (vonkajší kľukový hriadeľ) a jadra vankúša. Tu je tretí neurón vizuálneho analyzátora. Z nich sú vizuálne nervové vlákna poslané do kôry hemisférov veľkého mozgu.
Vo vonkajších (alebo bočných) kľukové hriadele, kde vlákna pochádzajú z sietnice, sú receptové polia, ktoré majú tiež zaoblený tvar, ale menej veľkosti ako v sietnici. Neuronov Reakcie Tu sú zariadenie, ale sú výraznejšie ako v sietnici.
Na úrovni vonkajších kľukových hriadeľov, proces interakcie aferentných signálov prichádzajúcich z sietnice oka, s účinnosťou oddelenia kortikálneho oddelenia vizuálneho analyzátora. Za účasti zverskej formácie je interakcia so sluchovými a inými senzorickými systémami, ktorý zabezpečuje procesy volebného zrýchlenia prostredníctvom pridelenia najvýznamnejších zložiek senzorického signálu.
Centrálne,alebo korok, oddelenievizuálny analyzátor sa nachádza v okcipitálnom zdieľaní (poliach 17, 18, 19 od Brodmana) alebo VI, V2, V3 (podľa prijatej nomenklatúry). Predpokladá sa, že primárna projekčná oblasť (pole 17) vykonáva špecializované, ale zložitejšie ako v sietnici a v externých kľukových hriadeľoch, spracovanie informácií. Receptové polia neurónov vizuálnej kôry malých veľkostí sú predĺžené, takmer obdĺžnikové a nie zaoblené formy. Spolu s týmto, existujú komplexné a kontrolované receptívne polia detektora typu. Táto funkcia vám umožňuje prideliť z pevného obrazu iba jednotlivé časti riadkov s rôznymi usporiadaniami a orientáciou, zatiaľ čo schopnosť selektívne reagovať na tieto fragmenty.
V každom segmente kortexu sa neuróny sústreďujú, že tvoriť stĺpec prebiehajúcej hĺbku cez všetky vrstvy vertikálne, zatiaľ čo funkčná asociácia neurónov, ktorí vykonávajú podobnú funkciu. Rôzne vlastnosti vizuálnych predmetov (farba, tvar, pohyb) sa spracúvajú v rôznych častiach vizuálneho kortexu veľkého mozgu paralelne.
Vizuálne cortex sú funkčne odlišné skupiny buniek - jednoduché a komplexné.
Jednoduché bunky vytvárajú receptívne pole, ktoré pozostáva z excitovaných a brzdových zón. To je možné určiť štúdiom bunkovej reakcie na malé svetlo. Štruktúra receptívneho poľa komplexnej bunky na vytvorenie tohto spôsobu je nemožné. Tieto bunky sú detektory uhla, naklápania a pohybu čiary v zornom poli.
V jednom stĺpci možno umiestniť jednoduché aj komplexné bunky. V vrstvách III a IV vizuálnej kôry, kde sa nachádzajú na konci thalamických vlákien, jednoduché bunky. Komplexné bunky sú umiestnené vo viacerých povrchových vrstvách poľa 17, v poliach 18 a 19 vizuálnej kôry, jednoduché bunky sú výnimkou, komplexné a ultra-prázdne bunky sa nachádzajú.
V vizuálnej kôre sa časť neurónov vytvorí "jednoduché" alebo sústredné farebné popisentné polia predpisu (IV vrstva). Farebná opponzácia RP sa prejavuje v tom, že Neuron, ktorý sa nachádza v strede, reaguje s excitáciou jednej farby a je inhibovaná počas stimulácie inej farby. Niektoré neuróny reagujú na respondentovi na červenom osvetlení a z reakcie na zelenú, reakcia druhých je opačná.
V neurónoch so sústredným RP, okrem oponentných vzťahov medzi farebnými reaktormi (Kolzkov), existujú antagonistické vzťahy medzi centrom a perifériou, t.j. Existujú RP s duálnou farbou popponencou. Napríklad, ak sa na expozícii v strede RP v Neurone nachádza na response na červenú a odozvu na zelenú, potom vyberie farbu na farbu s selektivitou na jas príslušnej farby, A nereaguje na difúznu stimuláciu svetelnými vlnami akejkoľvek dĺžky (z -puponentného vzťahu medzi stredom a perifériou RP).
V jednoduchom RP sa rozlišujú dva alebo tri paralelné zóny, medzi ktorými existujú dvojité oponenti: ak má centrálna zóna na-odozva na červené osvetlenie a odozvu na zelenú, potom okrajové zóny vydávajú odozvu na červenú a zapnutú -Retrote na zelenú.
Z ihriska VI - Ďalší (Dorzal) kanál prechádza cez sekundárne (mediotemapruzal - mt) oblasti kôry. Registrácia odpovedí neurónov tejto oblasti ukázala, že sú vysoko selektívne na jednorazovú (neidentitu), rýchlosť a smerovanie pohybu objektov sály, dobre reagujú na pohyb objektov na textúrovanom pozadí. Lokálne zničenie dramaticky zhoršuje schopnosť reagovať na pohybujúce sa objekty, ale po určitom čase sa táto schopnosť obnovuje, čo znamená, že táto oblasť nie je jedinou zónou, kde sa analýza pohyblivých objektov analyzuje v zornom poli. Ale spolu s týmto sa predpokladá, že informácie izolované neurónmi primárneho vizuálneho poľa 17 (V1) sa ďalej prenášajú na spracovanie do sekundárnej (pole V2) a terciárneho (pole V3) vizuálnej kôry.
Analýza vizuálnych informácií však nie je dokončená v poliach Striera (vizuálna) kôra (V1, V2, V3). Bolo zistené, že polia V1 začínajú (kanály) do iných oblastí, ktoré produkujú ďalšie spracovanie vizuálnych signálov.
Takže, ak zničíte pole V4 na opicu, ktorá je na križovatke časových a tmavých oblastí, potom je vnímanie farby a formy narušená. Spracovanie vizuálnych informácií o formulári, ako návrh, sa tiež vyskytuje najmä v nizhneirdial regióne. Pri zničení tejto oblasti, základné vlastnosti vnímania (zraková ostrosť a vnímanie svetla) netrpia, ale zlyhajú mechanizmy na analýzu najvyššej úrovne.
V vizuálnom zmyslovom systéme je teda komplikácia receptových polí neurónov z úrovne na úroveň, a tým vyššia je syntentifikovaná úroveň, strinst funkcie jednotlivých neurónov sú obmedzené.
V súčasnosti je vizuálny systém, počnúc gangliovými bunkami rozdelený na dve funkčne odlišné časti (magalové a parvocelulárne). Toto rozdelenie je spôsobené tým, že v cicavčích sietnici sú gangliové bunky rôznych typov - X, Y, W. Tieto bunky majú koncentrické receptívne polia a ich axóny tvoria vizuálne nervy.
V X-bunkách - RP je malý, s dobre vysloveným brzdovým okrajom, rýchlosť excitácie podľa ich osí - 15-25 m / s. RP je oveľa väčšie, sú lepšie odpovedať na difúzny lightware. Rýchlosť implementácie je 35-50 m / s. V renine X-bunky zaberajú centrálnu časť a ich hustota sa znižuje na okrajice. Y-bunky sú rovnomerne rozdelené na renine, preto na obvode sietnice je hustota Y buniek vyššia ako X bunky. Vlastnosti štruktúry RP X buniek určujú ich lepšiu reakciu na pomalý pohyb vizuálneho stimulu, zatiaľ čo y bunky reagujú lepšie na rýchle pohyblivé stimuly.
Retina tiež opisuje početnú skupinu W buniek. Toto sú najmenšie gangliové bunky, rýchlosť ich Axon - 5-9 m / s. Bunky tejto skupiny nie sú homogénne. Medzi nimi existujú bunky s koncentrickými a homogénnymi RP a bunkami, ktoré sú citlivé na pohyb stimulu cez pole recept. V rovnakej dobe, bunková reakcia nezávisí od smeru pohybu.
Separácia systému X, Y a W pokračuje na úrovni kľukového hriadeľa a vizuálnej kôry. Neuróny X majú fázový typ reakcie (aktivácia vo forme krátkeho blesku pulzov), ich receptívne polia sú prezentované v periférnych oblastiach pohľadu, latentné obdobie ich reakcie je menšie. Takáto sada vlastností ukazuje, že sú nadšené rýchlo sa pohybujúcimi anfentmi.
Neuróny X majú topický typ reakcie (Neuron je aktivovaný po dobu niekoľkých sekúnd), ich RPS sú prezentované vo väčšom počte v strede zorného poľa a latentné obdobie je väčšie.
Primárne a sekundárne zóny vizuálneho kortexu (polia Y1 a Y2) sa líšia v obsahu X a Y-neurónov. Napríklad v poli Y1 z vonkajšieho kľukového hriadeľa, affentácia pochádza z typu X aj z typu Y, zatiaľ čo pole Y2 prijíma afferety len z buniek typu Y.
Študovanie prenosu signálov na rôznych úrovniach systému vizuálneho snímača sa vykonáva registráciou celkových spôsobených potenciálov (VP) popredným osobou s pomocou elektród z povrchu kože hlavy v oblasti vizuálnej kôry (okcipitálna oblasť). U zvierat môžete súčasne preskúmať spôsobenú činnosť vo všetkých oddeleniach systému vizuálneho snímača.
Mechanizmy poskytujúce jasné vízie v rôznych podmienkach
Pri zohľadnení objektov umiestnených pri rôznych odstraňovaní od pozorovateľa, nasledujúce procesy prispievajú k jasným zrakom.
1. Konvergencia a odlišné pohyby očí,vďaka, ktorým sa vykoná zníženie alebo chov vizuálnych osí. Ak sa obe oči pohybujú v jednom smere, takéto pohyby sa nazývajú priateľské.
2. Reakcia žiakov,ktorý sa vyskytuje synchrónne s pohybom očí. Takže s konvergenciou vizuálnych osí, keď sa zvažujú úzke umiestnené objekty, žiak je zužovaný, t.j. konvergentná reakcia žiakov. Táto reakcia pomáha znížiť skreslenie obrazu spôsobeného sférickou aberáciou. Sférická aberácia je spôsobená tým, že refrakčné médiá očí majú nerovnaku ohniskovú vzdialenosť v rôznych oblastiach. Centrálna časť, cez ktorú prechádza optická os, má väčšiu ohnisko ako periférna časť. Preto sa obraz na sietnici získa necitlivý. Čím menší priemer žiaka, tým menšie skreslenie spôsobené sférickou aberáciou. Konvergentné zúženie žiaka zahŕňajú ubytovacie prístroje, ktoré spôsobuje zvýšenie refrakčnej sily šošovky.

Obr. 2.4. Mechanizmus ubytovania EYE: A - Mier, B - Napätie

Obr. 2.5
Žiak je tiež prístrojom eliminácie chromatickej aberácie, ktorá je spôsobená tým, že optické zariadenie oka, podobne ako jednoduché šošovky, refraktivuje svetlo s krátkou vlnou je silnejšia ako s dlhou vlnou. Na základe toho, pre presnejšie zameranie červenej farby, je potrebný veľký stupeň ubytovania ako pre modrú. To je dôvod, prečo sa modré objekty zdajú byť vzdialené ako červená, ktorá sa nachádza v rovnakej vzdialenosti.
3. Ubytovanie je hlavným mechanizmom, ktorý poskytuje jasnú víziu vyvinutých položiek a je znížená na zaostrovanie obrazu z diaľky alebo zatvorených položiek na sietnici. Hlavným mechanizmom ubytovania leží v nedobrovoľnej zmene v zakrivení objektívu oka (obr. 2.4).
Vďaka zmene zakrivenia šošovky, najmä predného povrchu, jej refrakčná sila sa môže líšiť v závislosti od 10-14 dioptrov. Kryštál sa uzatvára v kapsule, ktorý, pozdĺž okrajov (pozdĺž úrovne šošovky), ide do blokovacieho objektívu (Zinnov A Bunch), spojené s vláknami ciliárneho (ciliárneho) svalu. Pri znižovaní ciliárneho svalu klesá napätie zinnoy väzov a objektív vďaka svojej elasticitu sa stáva konvexným. Refrakčná sila sa zvyšuje a oko je nakonfigurované na víziu úzko usporiadaných položiek. Keď sa človek pozerá na diaľku, zväzok Qingnova je v natiahnutom stave, čo vedie k natiahnutiu kabelky objektívu a jeho zahusťovaniu. Innervation ciliárnych svalov sa vykonáva sympatickými a parasympatickými nervmi. Impulzia prechádza cez parasympatické vlákna ooo nervu spôsobuje svalovú kontrakciu. Sympatické vlákna odišli z najvyššej stránky krčka maternice, pretože relaxáciu. Zmena stupňa redukcie a relaxácie ciliárneho svalu je spojená s excitáciou sietnice a je pod vplyvom kortexu mozgu. Refrakčná sila oka je vyjadrená v dioptroch (E). Jedna dioptrová zodpovedá refrakčnej silu šošovky, ktorej hlavná ohnisková vzdialenosť je rovná 1 m. Ak je hlavnou ohniskovou vzdialenosťou šošovky napríklad, 0,5 alebo 2 m, potom je jeho refrakčná sila, resp. 2D alebo 0,5d. Refrakčná sila oka bez fenoménu ubytovania je 58-60 d a nazýva refrakcia oka.
S normálnym refrakciou očných lúčov z diaľkových položiek po prechode cez svetelný systém, oči sa zhromažďujú v zameraní na sietnicu v centrálnom YAM. Normálna refrakcia očí sa nazýva Emmetropi a také oko sa nazýva emmetropic. Spolu s normálnou refrakciou sa pozorovali jeho abnormality.
Myopia (Myopia) je taký typ znehodnotenia refrakcie, v ktorom sa lúče z predmetu po prechode cez svetelnú stroj nie sú zamerané na sietnicu, ale pred ním. Môže to závisieť od veľkej žiakovej sily oka alebo z vysokej dĺžky očnej buľvy. Zdá sa, že blízko ubytovania bez ubytovania, vzdialené položky vidí nejasné, nejasné. Na korekciu sa používajú okuliare s rozptyľovaním biconed šošoviek.
Hyperamerium (hyperopia) je forma porušenia refrakcie, v ktorej lúče z ďalekozrešných položiek kvôli slabému refrakčnej schopnosti oka alebo v malej dĺžke očnej kvality sa zameriavajú na sietnicu. Dokonca aj vzdialené predmety ďalekosiahlejšieho oka vidí so stresom ubytovania, v dôsledku čoho sa rozvíja hypertrofia ubytovaných svalov. Na korekciu sa používajú dvojité šošovky.
Astigmatizmus je forma znehodnotenia lomu, pri ktorom sa lúče nemôžu zbierať v jednom bode, v zameraní (z gréckeho. Stigme - bod), kvôli rôznym zakrivením rohovky a objektívu v rôznych meridiánoch (lietadlá). S astigmatizmom, objekty sa zdajú byť sploštené alebo predĺžené, jeho korekcia sa uskutočňuje sferocylindrickými šošovkami.
Treba poznamenať, že oko oka tiež zahŕňa: rohovka, vlhkosť prednej komory, krištáľ a sklovité telo. Avšak, ich refrakčná sila, na rozdiel od objektívu, nie je regulovaná a neprijíma účasť na ubytovaní. Po absolvovaní lúčov cez systém s refrakčným očným na sietnici sa vykazuje platný, redukovaný a obrátený obraz. Ale v procese individuálneho vývoja, porovnanie pocitov vizuálneho analyzátora s pocitmi motora, kože, vestibulárne a ďalších analyzátorov, ako je uvedené vyššie, vedie k tomu, že osoba vníma vonkajší svet, ako je to naozaj.
Binokulárne videnie (zrak s dvoma očami) hrá dôležitú úlohu pri vnímaní odvodených predmetov a určenie vzdialenosti k nim, dáva výraznejšie zmysel pre hĺbku priestoru v porovnaní s monokulárnou víziou, tj. víziu s jedným okom. Pri pohľade na položku s dvoma očami, jeho obraz môže padnúť na symetrické (identické) retinálne body oboch očí, excitácia, z ktorej sú kombinované v koncovom kortexu analyzátora na jedno celé číslo, čo dáva jeden obraz. Ak obraz objektu spadne na non-inscript (nevytečie) sietniciach, vystupuje rozdelenie obrazu. Proces vizuálnej analýzy priestoru závisí nielen na prítomnosti binokulárnej videnie, konvenčné a reflexné interakcie sa hrajú s významnou úlohou, ktorá sa vyvíja medzi vizuálnymi a motorickými analyzátormi. Následne sa nájdu konvergenčné pohyby oka a procesu ubytovania, ktoré sú spravované princípom spätnej väzby. Vnímanie priestoru ako celku je spojené s definíciou priestorových vzťahov viditeľných objektov - ich hodnoty, formulárov, vzťahov, ktoré je zabezpečené interakciou rôznych oddelení analyzátora; Významná úloha zohráva získaná skúsenosť.
Pri pohybujúcich sa objektochnasledujúce faktory prispievajú k jasnej vízii:
1) ľubovoľné pohyby oka smerom nahor, dole, vľavo alebo priamo pri rýchlosti pohybu objektu, ktorá sa vykonáva vďaka priateľským aktivitám okuliarov;
2) Keď sa v novom zornom poli zobrazí objekt, reflex fixácie sa spustí - rýchly nedobrovoľný pohyb oka, ktorý zaisťuje zarovnanie obrazu predmetu na sietnici s centrálnym vreckom. Pri sledovaní pohybujúceho sa objektu sa vyskytne pomalý pohyb oka - sledovací pohyb.
Pri zvažovaní pevného objektuaby sa zabezpečilo, že jasné oko zraku robí tri typy malých nedobrovoľných pohybov: Tremor - oko sa triasť s malou amplitou a frekvenciou, drift - pomalý posun oka na pomerne významnej vzdialenosti a skokoch (fliks) - rýchle pohyby očí. Tam sú tiež sakadické pohyby (Saccada) - priateľské pohyby oboch očí vykonaných pri vysokej rýchlosti. SACCADA je pozorovaná pri čítaní, prezeranie obrazov, keď pozorované body vizuálneho priestoru sú na jednej vzdialenosti od pozorovateľa a iných objektov. Ak blokujete tieto pohyby oka, svet okolo nás kvôli prispôsobeniu receptorov sietnice bude ťažké odlíšiť, ako je z žaby. Oči žaby sú stále, takže jasne rozlišuje len pohyblivé predmety, ako sú motýle. To je dôvod, prečo žaba sa blíži had, ktorý neustále hodí svoj jazyk. Had sa nerozlišuje v stave nehybnosti a jeho pohyblivý jazyk trvá na lietajúce motýľ.
V podmienkach zmeny svetlajasná vízia poskytuje reflexu žiakov, tmavú a ľahkú adaptáciu.
Zrenica Nastavuje intenzitu svetelného toku pôsobiaceho na sietnici zmenou jeho priemeru. Šírka žiaka sa môže pohybovať od 1,5 do 8,0 mm. Zúženie žiaka (MIOS) sa vyskytuje s rastúcim osvetľovaním, ako aj pri pohľade úzko umiestneným subjektom a vo sne. Rozšírenie žiaka (Midriaz) sa vyskytuje, keď sa osvetlenie znižuje, ako aj keď sú receptory nadšené, akékoľvek aferentné nervy, s emocionálnymi napäťovými reakciami spojenými so zvýšením tónu sympatickej jednotky nervového systému (bolesť, hnev , strach, radosť, atď.), S mentálnou excitáciou (psychóza, hystéria atď.), Pri udusení, anestézii. Pobiak Reflex Keď sa svetlo zmení, hoci zlepšuje vizuálne vnímanie (v tme je rozširovanie, čo zvyšuje prúdový prúd, padajúci na sietnicu, je zúžený do svetla), ale hlavný mechanizmus je stále tmavý a svetlý adaptácie.
Adaptácia Tempusje vyjadrená pri zvyšovaní citlivosti vizuálneho analyzátora (senzibilizácie), adaptácia svetla- pri znižovaní citlivosti oka na svetlo. Základom mechanizmov ľahkej a tmavej adaptácie sú fotochemické procesy prúdiajúce v tvarovanie a paličky, ktoré poskytujú rozdelenie (vo svetle) a resintez (v tme) fotosenzitívnych pigmentov, ako aj funkčné procesy mobility: zapnutie a vypnutie aktivity prvkov retinálneho receptora. Okrem toho, adaptácia definovať niektoré nervové mechanizmy a predovšetkým procesy vyskytujúce sa v nervových prvkoch sietnice, najmä spôsoby spájania fotoreceptorov na gangliové bunky zahŕňajúce horizontálne a bipolárne bunky. Počet receptorov spojených s jedným bipolárnym článkom sa teda zvyšuje v tme a väčšie ako ich číslo, ktoré sú na gangliovej bunke. V tomto prípade je receptové pole každého bipolárneho a prirodzene, gangliové bunky sa rozširuje, čo zlepšuje vizuálne vnímanie. Zahrnutie horizontálnych buniek je regulované CNS.
Zníženie tónu sympatického nervového systému (desilnelizácia oka) znižuje rýchlosť temnej adaptácie a podávanie adrenalínu má opačný účinok. Podráždenie retikulárnej tvorby mozgového drieku zvyšuje frekvenciu impulzov vo vláknach vizuálnych nervov. Účinok CNS na adaptívnych procesoch v sietnici sa tiež potvrdzuje skutočnosť, že citlivosť nešťastného oka sa zmení na svetlo pri osvetľovaní druhého oka a pod pôsobením zvuku, čuchových alebo chutných stimulov.
Adaptácia farieb.Najrýchlejšia a ostrá adaptácia (redukcia citlivosti) sa vyskytuje pod pôsobením modrého purpurového stimulu. Červený stimul zaberá priemernú pozíciu.
Okdeľovacie vnímanie veľkých predmetov a ich častiposkytuje centrálne a periférne vízie - zmeny v uhle pohľadu. Najčastejšie posúdenie malých položiek predmetu je zabezpečené v prípade, že obraz padá na žlté miesto, ktoré je lokalizované v centrálnom davom oka, ako v tomto prípade je najväčšia zraková ostrosť. Je to spôsobené tým, že v oblasti žltých škvŕn existujú iba stĺpce, ich najmenšie rozmery a každý kontakt s malým množstvom neurónov, čo zvyšuje vizuálnu ostrosť. Zvýšenie zraku je určená najmenším uhlom pohľadu, pod ktorou sú oči stále schopné vidieť dva body oddelene. Normálne oko je schopné rozlišovať medzi dvoma žiariacimi bodmi v uhle pohľadu v 1 ". Zvýšenie zraku takéhoto oka sa prijíma na jednotku. Pozricia ostrosť závisí od optických vlastností oka, konštrukčných prvkov sietnice a Prevádzka neuronálnych mechanizmov vodiča a centrálnych úsekov vizuálneho analyzátora. Stanovenie zrakovej ostrosti sa uskutočňuje pomocou písmena alebo rôznych typov štandardných tabuliek. Veľké objekty všeobecne a okolitý priestor sú vnímané hlavne z dôvodu periférnej vízie , poskytuje veľké zorné pole.
Zorné pole je priestor, ktorý možno vidieť pevným okom. K dispozícii je samostatné zorné pole ľavého a pravého oka, ako aj všeobecné pole pohľadu pre dve oči. Veľkosť oblasti vízie u ľudí závisí od hĺbky polohy očnej buľvy a tvaru abnormálneho oblúka a nosa. Hranice zorného poľa sú označené hodnotou uhla vytvoreného vizuálnou osou oka a lúčom sa vykonáva na extrémny viditeľný bod cez nodulárny bod oka do sietnice. Zorné pole nie je rovnaké v rôznych meridiánoch (smeroch). Kniha - 70 °, na poschodí - 60 °, prach - 90 °, Knutrics - 55 °. Achromatické pole pohľadu je väčšie ako chromatické vzhľadom na skutočnosť, že na obvode sietnice neexistujú žiadne receptory, ktoré vnímajú farbu (COLUMS). Zapnuté, farebné pole pohľadu nie je rovnaké pre rôzne farby. Najobľúbenejšie zorné pole pre zelenú, žltú, viac za červenú, ešte viac pre modrú. Hodnota zorného poľa sa líši v závislosti od osvetlenia. Achromatické pole pohľadu v súmraku sa zvyšuje, svetlo sa znižuje. Chromatické pole pohľadu, naopak, zvyšuje svetlo, znižuje sa za súmraku. Záleží na procese mobilizácie a demobilizácie fotoreceptorov (funkčná mobilita). S možnosťou Twilight Vision, zvýšenie počtu funkčných palíc, t.j. Ich mobilizácia vedie k zvýšeniu achrómu zorného poratu, zároveň k zníženiu počtu fungujúcich pleľôčok (ich demobilizácia) vedie k zníženiu chromatického zorného poľa (p.g. remintované).
Vizuálny analyzátor má tiež mechanizmus rozlišujúca dĺžka svetla -farebné videnie.
Farebné videnie, vizuálne kontrasty a po sebe idúce obrázky
Farebná vízia - schopnosť vizuálneho analyzátora reagovať na zmeny v dĺžke svetlej vlny s tvorbou pocitu farby. Určitá dĺžka vlny elektromagnetického žiarenia zodpovedá zmyslu pre určitú farbu. Pocit červenej farby teda zodpovedá pôsobeniu svetla s vlnovou dĺžkou 620-760 nm, a fialová - 390-450 nm, zostávajúce farby spektra majú medziproduktové parametre. Miešanie všetkých farieb dáva pocit bielej. V dôsledku zmiešania troch hlavných farieb spektra - červenej, zelenej, modrej fialovej - v inom pomere, môžete tiež získať vnímanie akýchkoľvek iných farieb. Cítia Farby sú spojené s osvetlením. Keď sa znižuje, červené farby sa najprv prestanú líšiť, neskôr - modrá. Vnímanie farieb je spôsobené hlavnými procesmi vyskytujúcimi sa vo fotoreceptoroch. Najväčším uznaním je trojzložková teória farby Lomonosov - Young - Helmholz-Lazareva, podľa ktorého existujú tri typy fotoreceptorov v sietnici - mesh, samostatne vnímajú červené, zelené a modré-fialové farby. Kombinácie excitácie rôznych plôch vedú k pocitu rôznych farieb a odtieňov. Jednotná excitácia troch typov colums dáva pocit bielej. Trojzložková teória farebného pohľadu dostala potvrdenie v elektrofyziologických štúdiách R. GRANITE (1947). Boli menom Modulátory, Kolkovka, ktoré boli nadšení pri zmene jasu svetla (štvrtý typ), boli nazývaní Dominators. Následne bola Metóda mikrospektúry schopná stanoviť, že aj jeden kolín môže absorbovať lúče rôznych vlnových dĺžok. Je to spôsobené prítomnosťou rôznych pigmentov citlivých na vlny rôznych dĺžok v každom plukovec.
Napriek presvedčivé argumenty trojzložkovej teórie vo fyziológii farby zobrazenia sú popísané fakty, ktoré nenájdu vysvetlenia z týchto pozícií. To umožnilo predložiť teóriu protivníkov alebo kontrastných, farieb, t.j. Vytvorte tzv. Oponent teóriu farebného pohľadu evald gering.
Podľa tejto teórie, v oku a / alebo v mozgu, existujú traja proces súpera: jeden - pre pocit červenej a zelenej, druhý - pre pocit žltej a modrej, tretí je kvalitatívne odlišný od prvého Dva procesy - pre čiernobielo. Táto teória je uplatniteľná na vysvetlenie prenosu informácií o farbe v nasledujúcich úsekoch vizuálneho systému: gangliové bunky sietnice, vonkajšie kľukové hriadele, kortikálne centrá pohľadov, kde farebné popponentné RP fungujú s ich stredom a perifériou.
Na základe získaných údajov sa teda môže predpokladať, že procesy v stĺpcoch viac zodpovedajú trojzložkovej teórii farby, zatiaľ čo pre nervové siete sietnice a prekrývajúce sa vizuálne centrá, teória kontrastných farieb BEREING je vhodný.
Pri vnímaní farby sa procesy prúdiaci v neurónoch rôznych úrovní vizuálneho analyzátora (vrátane sietnice) hrajú s určitou úlohou (vrátane sietnice), ktoré boli nazývané farebné popponentné neuróny. Pri pôsobení do oka žiarenia jednej časti spektra sú vzrušené a druhá brzdenie. Takéto neuróny sa podieľajú na kódovaní farieb.
Existujú anomálie farieb, ktoré sa môžu prejaviť vo forme čiastočnej alebo úplnej farebnej slepoty. Ľudia vo všeobecnosti nerozlišujúce farby sa nazývajú Agromat. Čiastočná farebná slepota prebieha v 8-10% mužov a 0,5% žien. Predpokladá sa, že farba filmu je spojená s nedostatkom určitých génov v pohlavnom nepáriacom chromozóme. Tri typy čiastočného farebného parku sa líšia: protandopia(Daltonizmus) - slepota hlavne na červenú. Tento typ farebnej komory bol prvýkrát opísaný v roku 1794 J. Daltonovým fyzikom, ktorý mal tento druh anomálie. Ľudia s takýmto druhom anomálie sa nazývajú "Krasnoslepi"; dateropia- Zníženie zeleného vnímania. Títo ľudia sa nazývajú "greenosleps"; titanopia- zriedka sa stretla s anomáliou. Zároveň ľudia vnímajú modré a fialové farby, nazývajú sa "fialové oddelené".
Z hľadiska trojzložkovej teórie farebného videnia je každý z typov anomálie výsledkom neprítomnosti jedného z troch colummer color-viditeľných substrátov. Pre diagnostiku poruchy pomoci farebnej pomoci, E. B. Rabin, ako aj špeciálne zariadenia, ktoré dostali názov anomaloskopy.Identifikácia rôznych farebných vizuálnych anomálie má veľký význam pri určovaní profesionálnej vhodnosti osoby pre rôzne druhy práce (vodič, pilot, umelec atď.).
Schopnosť odhadnúť dĺžku ľahkej vlny, ktorá sa prejavuje v schopnosti farbe, hrá významnú úlohu v ľudskom živote, čo ovplyvňuje emocionálnu sféru a aktivity rôznych telesných systémov. Červená farba spôsobuje pocit tepla, pôsobí vzrušujúce na psychiku, zvyšuje emócie, ale rýchlo pneumatiky, vedie k napätiu svalov, zvýšenie krvného tlaku, dýchania. Oranžová farba spôsobuje pocit zábavy a pohody, prispieva k tráveniu. Žltá farba vytvára dobrú, zvýšenú náladu, stimuluje víziu a nervový systém. Toto je najzábavnejšia farba. Zelená farba prevádzkuje osviežujúce a upokojujúce, užitočné v nespavosti, prepracovanie, znižuje krvný tlak, celkový tón telesa a je najpriaznivejší pre ľudí. Modrá farba spôsobuje pocit chladu a pôsobí na nervový systém upokojujúci, a silnejší ako zelená (obzvlášť priaznivá modrá farba pre ľudí so zvýšenou nervóznou excitovateľnosťou), viac ako so zelenou, znižuje krvný tlak a svalový tón. Fialová farba nie je toľko tak ako psychika uvoľňuje. Zdá sa, že ľudská psychika, po spektre červenej na fialku, prechádza celý rad emócií. Toto je založené na používaní lascherového testu na určenie emocionálneho stavu tela.
Veľkolepé kontrasty a konzistentné obrázky.Vizuálne pocity môžu pokračovať po ukončení podráždenia. Tento fenomén dostal názov po sebe nasledujúcich obrázkov. Veľkolepé kontrasty sú modifikované vnímanie dráždivosti v závislosti od okolitého svetla alebo farebného pozadia. Existujú koncepty svetlo a farebné vizuálne kontrasty. Fenomén kontrastu sa môže prejaviť v preháňaní skutočného rozdielu medzi dvoma simultánnymi alebo konzistentnými pocitmi, takže existujú simultánne a konzistentné kontrasty. Šedý pás na bielom pozadí sa zdá, že tmavší z rovnakého pásu sa nachádza na tmavom pozadí. Toto je príklad simultánneho kontrastu. Ak zvážime sivé na červenom pozadí, zdá sa, že je to nazelenalý, a ak si zvážime sivé na modrom pozadí, potom získa žltý odtieň. Toto je fenomén simultánneho farebného kontrastu. Kontrast sekvenčného farby je zmeniť farebný pocit pri prekladaní zobrazenia na bielom pozadí. Takže, ak sa pozriete na farbu maľovanú v červenej farbe, a potom preložíte oko na bielu, potom získava zelenú odtieň. Príčinou vizuálneho kontrastu je spôsoby, ktoré sa vykonávajú vo fotoreceptori a neuronálne prístroji sietnice. Základom je vzájomné brzdenie buniek týkajúcich sa rôznych retinálnych receptívnych polí a ich projekcie v kortikálnom oddelení analyzátorov.
- Anatómia pohľadu
Keď vedci vysvetľujú fenomén pohľadu Často porovnávajú oko s fotoaparátom. Svetlo, rovnako ako sa stáva so šošovkami zariadenia, vstupuje do oka cez malú dieru - žiak umiestnený v strede dúhovky. Žiak môže byť širší alebo už: Nastaví množstvo svetla. Ďalej sa svetlo posiela do zadnej steny oka - retina, v dôsledku čoho objaví určitý obraz v mozgu (obrázok, obraz). Rovnakým spôsobom, keď svetlo padá na zadnú stenu fotoaparátu, obraz je upevnený na filme.
Zvážte podrobnejšie, ako naša vízia funguje.
Po prvé, svetlo prijíma viditeľné časti oka, ku ktorému sa vzťahujú. Iris("Prihlásenie") a sclera (biela oka). Prejdite cez žiak, svetlo padá na objektív so zaostrením ( kryštál) Ľudské oko. Pod vplyvom svetla, žiak oko zužuje bez akéhokoľvek úsilia alebo ovládanie človeka. Je to preto, že jeden zo svalov IRIS - svižný - citlivé na svetlo a reaguje na to, rozširovanie. Zúženie žiaka je spôsobené automatickým riadením nášho mozgu. Moderné vlastné zameranie fotografických zariadení vytvárajú približne rovnakú vec: fotoelektrické "oko" reguluje priemer vstupu za šošovkou, čím sa dávkovanie množstva svetla.
Teraz sa poďme na miesto ležiace za oku, kde sa nachádza objektív, sklovitý Cente ( tela) A nakoniec - retina, Orgán, ktorý spôsobuje originálny obdiv na jeho štruktúru. Retina pokrýva rozsiahly povrch oka. Ide o jedinečný orgán s komplexnou štruktúrou, ktorá nemá rád žiadnu inú štruktúru tela. Shell shell oka pozostáva zo stoviek miliónov fotosenzitívnych buniek, nazývaných "paličky" a "Kolkoks". nereformované svetlo. Palice Navrhnuté tak, aby videli v tme a keď sú zapojené, môžeme vnímať neviditeľné. Film nie je schopný. Ak používate film určený na natáčanie v súmraku, nebude schopný zachytiť obraz viditeľný počas jasného svetla. Ľudské oko však má len jednu sietnicu a je schopná pôsobiť v rôznych podmienkach. Možno sa môže nazvať polyfunkčným filmom. StĺpceNa rozdiel od paličky, pracujte najlepšie, keď svetlo. Potrebujú svetlo, aby poskytli jasné zaostrenie a čistý zrak. Najvyššia koncentrácia hrubého čreva - v oblasti sietnice, ktorá sa nazýva Makula ("škvrna"). V centrálnej časti tejto farby sa nachádza Fovea Centralis: je to táto oblasť, ktorá umožňuje najviac akútnej vízie.
Rohovka, žiak, kryštál, sklovité telo, podobne ako veľkosť oka jablko, závisí od zaostrenia svetla, pretože prechádza cez tie alebo iné štruktúry. Proces zmeny zamerania svetla sa nazýva lom (refrakcia). Svetlo zamerané presnejšie padá na fob, zatiaľ čo menej zamerané svetlo sa rozptyľuje na sietnici.
Naše oči sú schopní rozlišovať medzi desiatimi miliónmi stupňov intenzity svetla a asi sedem miliónov odtieňov farieb.
Avšak, anatómia vízie sa na to nevohraná. Osoba, ktorú vidí, zároveň používa oči a mozog, a to nie je dosť jednoduchej analógie s fotoaparátom. Každú sekundu oko posiela do mozgu o milión jednotiek informácií (viac ako 75% informácií vnímaných nami). Tieto časti svetla sa zameriavajú na vedomie prekvapivo zložitým obrazom, ktoré identifikujete. Svetlo, pričom tvar týchto rozpoznateľných obrázkov sa javí ako zvláštny stimulant pre vaše spomienky na minulé udalosti. V tomto zmysle vízia pôsobí len ako pasívne vnímanie.
Takmer všetko, čo vidíme, je to, čo sme sa naučili vidieť. Koniec koncov, dostávame k životu bez toho, aby ste mali nápady, ako produkovať informácie zo svetla padajúceho na sietnicu. V detstve, čo vidíme pre nás neznamená nič alebo takmer nič. Svetlým stimulovaným impulzom z sietnice spadajú do mozgu, ale sú pre dieťa - len pocit, ktoré nemajú význam. Ako vyrastáme a učíme sa, osoba začína interpretovať tieto pocity, snažiť sa ich prísť, pochopiť, čo znamenajú.
| Články na tému: | |
|
Hydridový systém skladovania vodíka rozdiely fenolov z alkoholov
Lopta a perník železná graf (i) hydrid (FEH) molekula .... Katedra koloidnej chémie
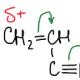
hlava Katedra chemických vied, senior výskumný pracovník ... Charakteristické chemické vlastnosti uhľovodíkov
Vytvára sa pri prekrývaní atómových orbitálov a tvorby spoločných ... | |