вибір читачів

Популярні статті
синапс
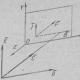
Як передається збудження від одного нейрона іншому або від нейрона, наприклад, на м'язове волокно? Цією проблемою цікавляться не тільки професійні нейробіологи, а й лікарі, особливо фармакологи. Знання біологічних механізмів необхідно для лікування деяких захворювань, а також для створення нових ліків і препаратів. Справа в тому, що одними з основних місць дії цих речовин на організм людини є місця передачі збудження з одного нейрона на інший (або на іншу клітину, наприклад клітку серцевого м'яза, стінки судин і ін.). Відросток нейрона аксон направляється до іншого нейрона і утворює на ньому контакт, який називають синапсом(В перекладі з грецького - контакт; див. Рис. 2.3). Саме синапс зберігає багато таємниць мозку. Порушення цього контакту, наприклад, речовинами, які блокують його роботу, призводить до важких наслідків для людини. Це місце для вкладання дії наркотиків. Приклади будуть приведені нижче, а зараз розглянемо, як влаштований і як працює синапс.
Труднощі цього дослідження визначаються тим, що сам синапс дуже маленький (його діаметр не більше 1 мкм). Один нейрон отримує такі контакти, як правило, від декількох тисяч (3 - 10 тис.) Інших нейронів. Кожен синапс надійно закритий спеціальними клітинами глії, тому досліджувати його дуже непросто. На рис. 2.12 показана схема синапсу, як це уявляє собі сучасна наука. Незважаючи на свою мініатюрність, він влаштований досить складно. Одним з його основних компонентів є бульбашки, які знаходяться всередині синапсу. Ці бульбашки містять біологічно дуже активна речовина, яка називається нейротрансмиттером, або медіатором (Передавачем).
Згадаймо, що нервовий імпульс (збудження) з величезною швидкістю просувається по волокну і підходить до синапси. Цей потенціал дії викликає деполяризацію мембрани синапсу (рис. 2.13), однак це не призводить до генерації нового порушення (потенціалу дії), а викликає відкривання спеціальних іонних каналів, з якими ми ще не знайомі. Ці канали пропускають іони кальцію всередину синапсу. Іони кальцію відіграють дуже велику роль в діяльності організму. Спеціальна залоза внутрішньої секреції - паращитовидная (вона знаходиться поверх щитовидної залози) регулює вміст кальцію в організмі. Багато захворювань пов'язані з порушенням обміну кальцію в організмі. Наприклад, його нестача призводить до рахіту у маленьких дітей.
Яким чином кальцій бере участь в роботі синапсу? Потрапляючи в цитоплазму синаптического закінчення, кальцій входить в зв'язок з білками, утворюють оболонку бульбашок, в яких зберігається медіатор. В кінцевому підсумку мембрани синаптичних бульбашок стискаються, виштовхуючи свій вміст в синаптичну щілину. Цей процес дуже нагадує скорочення м'язового волокна в м'язі, в усякому разі, ці два процеси мають однаковий механізм на молекулярному рівні. Таким чином, зв'язування кальцію білками оболонки бульбашки призводить до її скорочення, і зміст бульбашки впорскується (екзоцитоз) в щілину, яка відокремлює мембрану одного нейрона від мембрани іншого. Ця щілина називається синоптичної щілиною. З опису має бути ясно, що збудження (електричний потенціал дії) нейрона в синапсі перетворюється з електричного імпульсу в імпульс хімічний. Іншими словами, кожне порушення нейрона супроводжується викидом в закінченні його аксона порції біологічно активної речовини - медіатора. Далі молекули медіатора зв'язуються з спеціальними білковими молекулами, які перебувають на мембрані іншого нейрона. Ці молекули називаються рецепторами. Рецептори влаштовані унікально і пов'язують тільки один тип молекул. У деяких описах вказується, що вони підходять, як «ключ до замка» (ключ підходить тільки до свого замку).
Рецептор складається з двох частин. Одну можна назвати «який дізнається центром», іншу - «іонним каналом». Якщо молекули медіатора зайняли певні місця (довідається центр) на молекулі рецептора, то іонний канал відкривається і іони починають входити в клітку (іони натрію) або виходити (іони калію) з клітки. Іншими словами, через мембрану протікає іонний струм, який викликає зміну потенціалу на мембрані. Цей потенціал отримав назву постсинаптичного потенціалу (Рис. 2.13). Дуже важливою властивістю описаних іонних каналів є те, що кількість відкритих каналів визначається кількістю пов'язаних молекул медіатора, а не потенціалом на мембрані, як у випадку з Електровозбудімость мембраною нервового волокна. Таким чином, постсинаптические потенціали мають властивість градуально: амплітуда потенціалу визначається кількістю молекул медіатора, пов'язаного рецепторами. Завдяки цій залежності амплітуда потенціалу на мембрані нейрона розвивається пропорційно кількості відкритих каналів.
На мембрані одного нейрона можуть одночасно перебувати два види синапсів: гальмівні і збудливі. Все визначається пристроєм іонного каналу мембрани. Мембрана збудливих синапсів пропускає як іони натрію, так і іони калію. В цьому випадку мембрана нейрона деполяризуется. Мембрана гальмівних синапсів пропускає тільки іони хлору і гиперполяризуется. Очевидно, що якщо нейрон загальмований, потенціал мембрани збільшується (гиперполяризация). Таким чином, нейрон завдяки впливу через відповідні синапси може збудитися або припинити порушення, загальмуватися. Всі ці події відбуваються на сома і численних відростках дендрита нейрона, на останніх знаходиться до декількох тисяч гальмівних і збудливих синапсів.
Як приклад розберемо, як діє в синапсі медіатор, який називається ацетилхолином. Цей медіатор широко поширений в головному мозку і в периферичних закінченнях нервових волокон. Наприклад, рухові імпульси, які за відповідними нервах призводять до скорочення м'язів нашого тіла, оперують ацетилхолином. Ацетилхолін був відкритий в 30-х роках австрійським вченим О. Леві. Експеримент був дуже простий: ізолювали серце жаби з відповідним до нього блукаючим нервом. Було відомо, що електрична стимуляція блукаючого нерва призводить до уповільнення скорочень серця аж до повної його зупинки. О. Леві простимулював блукаючий нерв, отримав ефект зупинки серця і взяв з серця трохи крові. Виявилося, що якщо цю кров додати в шлуночок працюючого серця, то воно уповільнює свої скорочення. Був зроблений висновок: при стимуляції блукаючого нерва виділяється речовина, яка зупиняє серце. Це і був ацетилхолін. Пізніше був відкритий фермент, який розщеплював ацетилхолін на холін (жир) і оцтову кислоту, в результаті чого припинялася дія медіатора. Цим дослідженням вперше була встановлена \u200b\u200bточна хімічна формула медіатора і послідовність подій в типовому хімічному синапсі. Ця послідовність подій зводиться до наступного.
Потенціал дії, що прийшов по пресинаптичними волокну до синапси, викликає деполяризацію, яка включає кальцієвий насос, і іони кальцію надходять в синапс; іони кальцію зв'язуються білками мембрани синаптичних пухирців, що призводить до активного спорожнення (екзоцитозу) бульбашок в синаптичну щілину. Молекули медіатора зв'язуються (який дізнається центром) відповідними рецепторами постсинаптичної мембрани, при цьому відкривається іонний канал. Через мембрану починає протікати іонний струм, що призводить до виникнення на ній постсинаптичного потенціалу. Залежно від характеру відкритих іонних каналів виникає збудливий (відкриваються канали для іонів натрію і калію) або гальмівний (відкриваються канали для іонів хлору) постсинаптичний потенціал.
Ацетилхолін вельми широко поширений в живій природі. Наприклад, він знаходиться в жалких капсулах кропиви, в жалких клітинах кишковопорожнинних тварин (наприклад, прісноводної гідри, медузи) та ін. У нашому організмі ацетилхолін викидається в закінченнях рухових нервів, керуючих м'язами, з закінчень блукаючого нерва, який керує діяльністю серця і інших внутрішніх органів. Людина давно знайомий з антагоністом ацетилхоліну - це отрута кураре, яким користувалися індіанці Південної Америки при полюванні на тварин. Виявилося, що кураре, потрапляючи в кров, викликає знерухомлення тварини, і воно гине фактично від задухи, але кураре не зупиняє серце. Дослідження показали, що в організмі існують два типи рецепторів до ацетилхоліну: один успішно пов'язує нікотинову кислоту, а інший - мускарин (речовина, яка виділена з гриба роду Muscaris). На м'язах нашого тіла знаходяться рецептори нікотинового типу до ацетилхоліну, тоді як на серцевому м'язі і нейронах головного мозку - рецептори до ацетилхоліну мускаринового типу.
В даний час в медицині широко застосовують синтетичні аналоги кураре для знерухомлення хворих під час складних операцій на внутрішніх органах. Застосування цих засобів призводить до повного паралічу рухової мускулатури (зв'язується рецепторами нікотинового типу), але не впливає на роботу внутрішніх оранів, в тому числі серця (рецептори мускаринового типу). Нейрони головного мозку, порушувані через мускаринові ацетилхолінові рецептори, відіграють велику роль в прояві деяких психічних функцій. Зараз відомо, що загибель таких нейронів призводить до старечого слабоумства (хвороба Альцгеймера). Іншим прикладом, який повинен показати важливість саме рецепторів нікотинового типу на м'язі до ацетилхоліну, може служити захворювання, зване miastenia grevis (м'язова слабкість). Це генетично успадкована хвороба, т. Е. Її походження пов'язане з «поломками» генетичного апарату, які передаються у спадок. Захворювання проявляється у віці ближче до статевої зрілості і починається з м'язової слабкості, яка поступово посилюється і захоплює все більш великі групи м'язів. Причиною цієї недуги виявилося те, що організм хворого виробляє білкові молекули, які прекрасно зв'язуються ацетилхолінових рецепторів нікотинового типу. Займаючи ці рецептори, вони перешкоджають зв'язуванню з ними молекул ацетилхоліну, що викидаються з синаптичних закінчень рухових нервів. Це і призводить до блокування синаптического проведення до м'язів і, отже, до їх паралічу.
Описаний на прикладі ацетилхоліну тип синаптичної передачі - не єдиний в ЦНС. Другий тип синаптичноїпередачі також широко поширений, наприклад, в синапсах, в яких медіаторами є біогенні аміни (дофамін, серотонін, адреналін та ін.). У цьому типі синапсів має місце наступна послідовність подій. Після того як утворився комплекс «молекула медіатора - рецепторний білок», активується спеціальний мембранний білок (G-білок). Одна молекула медіатора при зв'язуванні з рецептором може активувати багато молекул G-білка, і це підсилює ефект медіатора. Кожна активована молекула G-білка в одних нейронах може відкривати іонний канал, а в інших активувати всередині клітини синтез спеціальних молекул, так званих вторинних посередників. Вторинні посередники можуть запускати в клітці багато біохімічні реакції, пов'язані з синтезом, наприклад, білка, в цьому випадку виникнення електричного потенціалу на мембрані нейрона не відбувається.
Існують і інші медіатори. У головному мозку в якості медіаторів «працює» ціла група речовин, які об'єднані під назвою біогенні аміни. В середині минулого століття англійський лікар Паркінсон описав хворобу, яка проявлялася як тремтливий параліч. Це важке страждання викликано руйнуванням в мозку хворого нейронів, які в своїх синапсах (закінченнях) виділяють дофамін -речовина з групи біогенних амінів. Тіла цих нейронів знаходяться в середньому мозку, утворюючи там скупчення, яке називається чорної субстанцією.Дослідження останніх років показали, що дофамін в мозку ссавців також має кілька типів рецепторів (в даний час відомо шість типів). Інша речовина з групи біогенних амінів - серотонін (інша назва 5-оксітріптамін) - спочатку було відомо як засіб, що приводить до підйому кров'яного тиску (судинозвужувальну). Зверніть увагу, що, це відображено в його назві. Однак виявилося, що виснаження в головному мозку серотоніну призводить до хронічного безсоння. У дослідах на тваринах було встановлено, що руйнування в мозковому стовбурі (задніх відділах мозку) спеціальних ядер, які відомі в анатомії як ядра шва, призводить до хронічного безсоння і надалі загибелі цих тварин. Біохімічне дослідження встановило, що нейрони ядер шва містять серотонін. У пацієнтів, які страждають хронічним безсонням, також виявлено зниження концентрації серотоніну в мозку.
До біогенних амінів відносять також адреналін і норадреналін, які містяться в синапсах нейронів автономної нервової вегетативної системи. Під час стресу під впливом спеціального гормону - адренокортикотропного (докладніше див. Нижче) - з клітин кори надниркових залоз в кров також викидаються адреналін і норадреналін.
З вищевикладеного зрозуміло, яке значення у функціях нервової системи відіграють медіатори. У відповідь на прихід нервового імпульсу до синапси відбувається викид медіатора; молекули медіатора з'єднуються (комплементарно - як «ключ до замка») з рецепторами постсинаптичної мембрани, що призводить до відкривання іонного каналу або до активування внутрішньоклітинних реакцій. Приклади синаптичної передачі, розглянуті вище, повністю відповідають цій схемі. Разом з тим завдяки дослідженням останніх десятиліть ця досить проста схема хімічної синаптичної передачі значно ускладнилася. Поява иммунохимических методів дозволило показати, що в одному синапсі можуть співіснувати кілька груп медіаторів, а не один, як це припускали раніше. Наприклад, в одному синаптическом закінчення одночасно можуть перебувати синаптичні пухирці, що містять ацетилхолін і норадреналін, які досить легко ідентифікуються на електронних фотографіях (ацетилхолін міститься в прозорих бульбашках діаметром близько 50 нм, а норадреналін - в електронно-щільних діаметром до 200 нм). Крім класичних медіаторів, в синаптическом закінчення можуть перебувати один або кілька ній-ропептідов. Кількість речовин, що містяться в синапсі, може доходити до 5-6 (своєрідний коктейль). Більш того, медіаторная специфічність синапсу може змінюватися в онтогенезі. Наприклад, нейрони симпатичних гангліїв, що іннервують потові залози у ссавців, початково норадренергічни, але у дорослих тварин стають холінергічних.
В даний час при класифікації медіаторних речовин прийнято виділяти: первинні медіатори, супутні медіатори, медіатори-модулятори і аллостерічеськіє медіатори. Первинними медіаторами вважають ті, які діють безпосередньо на рецептори постсинаптичної мембрани. Супутні медіатори і медіатори-модулятори можуть запускати каскад ферментативних реакцій, які, наприклад, фосфорилируют рецептор для первинного медіатора. Аллостерічеськіє медіатори можуть брати участь в кооперативних процесах взаємодії з рецепторами первинного медіатора.
Довгий час за зразок брали синаптичну передачу по анатомічній адресою (принцип «точка - в точку»). Відкриття останніх десятиліть, особливо медиаторной функції нейропептидів, показали, що в нервовій системі можливий принцип передачі і за хімічним адресою. Іншими словами, медіатор, що виділяється з даного закінчення, може діяти не тільки на «свою» постсинаптическую мембрану, а й за межами даного синапсу - на мембрани інших нейронів, що мають відповідні рецептори. Таким чином, фізіологічна реакція забезпечується не точним анатомічним контактом, а наявністю відповідного рецептора на клітці-мішені. Власне цей принцип був давно відомий в ендокринології, а дослідження останніх років знайшли йому більш широке застосування.
Всі відомі типи хеморецепторів на постсинаптичні мембрані поділяють на дві групи. В одну групу входять рецептори, до складу яких включено іонний канал, що відкривається при зв'язуванні молекул медіатора з «який дізнається» центром. Рецептори другої групи (метаботропние рецептори) відкривають іонний канал опосередковано (через ланцюжок біохімічних реакцій), зокрема, за допомогою активації спеціальних внутрішньоклітинних білків.
Одними з найпоширеніших є медіатори, що належать до групи біогенних амінів. Ця група медіаторів досить надійно ідентифікується мікрогістологіческімі методами. Відомі дві групи біогенних амінів: катехоламіни (дофамін, норадреналін і адреналін) і індоламін (серотонін). Функції біогенних амінів в організмі дуже різноманітні: медіаторная, гормональна, регуляція ембріогенезу.
Основним джерелом норадренергических аксонів є нейрони блакитного плями і прилеглих ділянок середнього мозку (рис. 2.14). Аксони цих нейронів широко поширюються в мозковому стовбурі, мозочку, в великих півкулях. У довгастому мозку велике скупчення норадренергических нейронів знаходиться в вентролатеральном ядрі ретикулярної формації. У проміжному мозку (гіпоталамусі) норадренергические нейрони поряд з дофаминергическими нейронами входять до складу гіпоталамо-гіпофізарної системи. Норадренергические нейрони в великій кількості містяться в периферичної нервової системи. Їх тіла лежать в симпатичної ланцюжку і в деяких інтрамуральних гангліях.
Дофамінергічні нейрони у ссавців знаходяться переважно в середньому мозку (так звана нігро-неостріарная система), а також в гіпоталамічної області. Дофамінові ланцюга мозку ссавців добре вивчені. Відомі три головні ланцюги, всі вони складаються з однонейронной ланцюжка. Тіла нейронів знаходяться в мозковому стовбурі і відсилають аксони в інші області головного мозку (рис. 2.15).
Одна ланцюг дуже проста. Тіло нейрона знаходиться в області гіпоталамуса і відсилає короткий аксон в гіпофіз. Цей шлях входить до складу гіпоталамо-гіпофізарної системи і контролює систему ендокринних залоз.
Друга дофаминовая система також добре вивчена. Це чорна субстанція, багато клітини якої містять дофамін. Аксони цих нейронів проектуються в смугасті тіла. Ця система містить приблизно 3/4 дофаміну головного мозку. Вона має вирішальне значення в регулюванні тонічних рухів. Дефіцит дофаміну в цій системі призводить до хвороби Паркінсона. Відомо, що при цьому захворюванні відбувається загибель нейронів чорної субстанції. Введення L-DOPA (попередника дофаміну) полегшує у хворих деякі симптоми захворювання.
Третя дофаминергическая система бере участь в прояві шизофренії та деяких інших психічних захворювань. Функції цієї системи поки вивчені недостатньо, хоча самі шляхи добре відомі. Тіла нейронів лежать в середньому мозку поряд з чорною субстанцією. Вони проектують аксони в вищерозміщені структури мозку, мозкову кору і лімбічну систему, особливо до фронтальній корі, до септальних області і енторінальной корі. Енторінальная кора, в свою чергу, є головним джерелом проекцій до гиппокампу.
Згідно дофаминовой гіпотезі шизофренії, третя дофаминергическая система при цьому захворюванні надактивного. Ці уявлення виникли після відкриття речовин, що знімають деякі симптоми захворювання. Наприклад, хлорпромазин і галоперидол мають різну хімічну природу, але вони однаково пригнічують активність дофаминергической системи мозку і прояв деякі симптомів шизофренії. У хворих на шизофренію, протягом року отримували ці препарати, з'являються рухові порушення, що отримали назву tardive dyskinesia (повторювані химерні руху лицьової мускулатури, включаючи мускулатуру рота, які хворий не може контролювати).
Серотонін майже одночасно відкрили в якості сироваткового сосудосуживающего фактора (1948) і ентераміна, що секретується ентерохромаффіновимі клітинами слизової оболонки кишечника. У 1951 р було розшифровано хімічну будову серотоніну і він отримав нову назву - 5-гідрокси. В організмі ссавців він утворюється гідроксилюванням амінокислоти триптофану з подальшим декарбоксилюванням. 90% серотоніну утворюється в організмі ентерохромаффіновимі клітинами слизової оболонки всього травного тракту. Внутрішньоклітинний серотонін інактивується моноаміноксидазою, що міститься в мітохондріях. Серотонін позаклітинного простору окислюється перулоплазміном. Велика частина виробленого серотоніну зв'язується з кров'яними пластинками і по кров'яному руслу розноситься по організму. Інша частина діє в якості місцевого гормону, сприяючи авторегулирования кишкової перистальтики, а також модулюючи епітеліальну секрецію і всмоктування в кишковому тракті.
Серотонінергічні нейрони широко поширені в центральній нервовій системі (рис. 2.16). Вони виявляються в складі дорсального і медіального ядер шва довгастого мозку, а також в середньому мозку і вароліевом мосту. Серотонінергічні нейрони іннервують великі ділянки мозку, що включають кору великих півкуль, гіпокамп, блідий шар, мигдалину, область гіпоталамуса. Інтерес до серотоніну був притягнутий в зв'язку з проблемою сну. При руйнуванні ядер шва тварини страждали безсонням. Подібний ефект надавали речовини, що виснажують сховище серотоніну в мозку.
Найвища концентрація серотоніну виявлена \u200b\u200bв епіфізі (pineal gland). Серотонін в епіфізі перетворюється в мелатонін, який бере участь в пігментації шкіри, а також впливає у багатьох тварин на активність жіночих гонад. Зміст як серотоніну, так і мелатоніну в епіфізі контролюється циклом світло - темрява через нервову симпатичну систему.
Іншу групу медіаторів ЦНС складають амінокислоти. Вже давно відомо, що нервова тканина з її високим рівнем метаболізму містить значні концентрації цілого набору амінокислот (перераховані в порядку убування): глутамінової кислоти, глутаміну, аспарагінової кислоти, гамма-аміномасляної кислоти (ГАМК).
Глутамат в нервовій тканині утворюється переважно з глюкози. У ссавців найбільше глутамату міститься в кінцевому мозку і мозочку, де його концентрація приблизно в 2 рази вище, ніж в стовбурі мозку і спинному мозку. У спинному мозку глутамат розподілено нерівномірно: в задніх рогах він знаходиться в більшій концентрації, ніж в передніх. Глутамат є одним з найпоширеніших медіаторів в ЦНС.
Постсинаптические рецептори до глутамату класифікуються відповідно до аффинностью (спорідненістю) до трьох екзогенних агонистам - квісгулату, каїнатом і N-метил-D-аспартату (NMDA). Іонні канали, що активуються квісгулатом і каїнатом, подібні каналам, які управляються нікотиновими рецепторами - вони пропускають суміш катіонів (Na + і. До +). Стимуляція NMDA-рецепторів має складний характер активації: іонний струм, який переноситься не тільки Na + і К +, але також Са ++ при відкриванні іонного каналу рецептора, залежить від потенціалу мембрани. Вольтзавісімая природа цього каналу визначається різним ступенем його блокування іонами Mg ++ з урахуванням рівня мембранного потенціалу. При потенціалі спокою порядку - 75 мВ іони Mg ++, які переважно знаходяться в міжклітинному середовищі, конкурують з іонами Са ++ і Na + за відповідні канали мембрани (рис. 2.17). Внаслідок того, що іон Mg ++ не може пройти через пору, канал блокується щоразу, як потрапляє туди іон Mg ++. Це призводить до зменшення часу відкритого каналу і провідності мембрани. Якщо мембрану нейрона деполярізовать, то кількість іонів Mg ++, які закривають іонний канал, знижується і через канал безперешкодно можуть проходити іони Са ++, Na + і. До +. При рідкісних стимуляції (потенціал спокою змінюється мало) глутаматергіческіх рецептораВПСП виникає переважно за рахунок активації квісгулатних і каінатного рецепторів; внесок NMDA-рецепторів незначний. При тривалій деполяризації мембрани (ритмічна стимуляція) магнієвий блок видаляється, і NMDA-канали починають проводити іони Са ++, Na + і. До +. Іони Са ++ через вторинні посередники можуть потенціювати (посилювати) мінПСП, що може привести, наприклад, до тривалого збільшення синаптичної провідності, що зберігається годинами і навіть цілодобово.
З гальмівних медіаторів ГАМК є найпоширенішою в ЦНС. Вона синтезується з L-глутамінової кислоти в одну стадію ферментом декарбоксилаз, наявність якої є лімітуючим фактором цього медіатора. Відомо два типи ГАМК-рецепторів на постсинаптичні мембрані: ГАМКА (відкриває канали для іонів хлору) і ГАМКб (відкриває в залежності від типу клітини канали для. До + або Са ++). На рис. 2.18 показана схема ГАМК-рецептора. Цікаво, що в його склад входить бензодіазіпіновий рецептор, наявністю якого пояснюють дію так званих малих (денних) транквілізаторів (седуксен, тазепама і ін.). Припинення дії медіатора в ГАМК-синапсах відбувається за принципом зворотного всмоктування (молекули медіатора спеціальним механізмом поглинаються з синаптичної щілини в цитоплазму нейрона). З антагоністів ГАМК добре відомий бікукулін. Він добре проходить через гематоенцефалічний бар'єр, робить сильний вплив на організм навіть в малих дозах, викликаючи конвульсії і смерть. ГАМК виявляється в ряді нейронів мозочка (в клітинах Пуркіньє, клітинах Гольджі, корзинчатих клітинах), гіпокампу (в корзинчатих клітинах), в нюхової цибулини і чорної субстанції.
Ідентифікація ГАМК-ланцюгів мозку важка, так як ГАМК - звичайний учасник метаболізму в ряді тканин організму. Метаболічна ГАМК не використовується як медіатор, хоча в хімічному відношенні їх молекули однакові. ГАМК визначається по ферменту декарбоксилази. Метод заснований на отриманні у тварин антитіл до декарбоксилази (антитіла екстрагують, мітять і вводять в мозок, де вони зв'язуються з декарбоксилаз).
Іншим відомим гальмівним медіатором є гліцин. Гліцинергічними нейрони знаходяться головним чином в спинному і довгастому мозку. Вважають, що ці клітини виконують роль гальмівних интернейронов.
Ацетилхолін - один з перших вивчених медіаторів. Він надзвичайно широко поширений в периферичної нервової системи. Прикладом можуть служити мотонейрони спинного мозку і нейрони ядер черепних нервів. Як правило, холинергические ланцюга в мозку визначають по присутності ферменту холінестерази. У головному мозку тіла холинергических нейронів знаходяться в ядрі перегородки, ядрі діагонального пучка (Брока) і базальних ядрах. Нейроанатомии вважають, що ці групи нейронів формують фактично одну популяцію холинергических нейронів: ядро \u200b\u200bпеднего мозку, nucleus basalis (воно розташоване в базальної частини переднього мозку) (рис. 2.19). Аксони відповідних нейронів проектуються до структур переднього мозку, особливо в нову кору і гіпокамп. Тут зустрічаються обидва типи ацетилхолінових рецепторів (мускаринові і нікотинові), хоча вважається, що мускаринові рецептори домінують в більш рострально распоженних мозкових структурах. За даними останніх років складається враження, що ацетилхолінового система відіграє велику роль в процесах, пов'язаних з вищими інтегративними функціями, які вимагають участі пам'яті. Наприклад, показано, що в мозку хворих, які померли від хвороби Альцгеймера, спостерігається масивна втрата холінергічних нейронів в nucleus basalis.
Нервові клітини керують функціями організму за допомогою хімічних сигнальних речовин, нейромедіаторів і нейрогормонів. нейромедіатори - короткоживучі речовини локальної дії; вони виділяються в синаптичну щілину і передають сигнал сусіднім клітинам (продукуються нейронами і зберігаються в синапсах; при надходженні нервового імпульсу вони виділяються в синаптичну щілину, вибірково зв'язуються з спеціфіческімрецептором на постсинаптичні мембрані іншого нейрона або м'язової клітини, стимулюючи ці клітини до виконання ними своїх специфічних функцій). Речовина, з якого синтезується медіатор (попередник медіатора), потрапляє в нейрон або його закінчення з крові або ліквору (рідина, що циркулює в головному і спинному мозку) і в результаті біохімічних реакцій під впливом ферментів перетворюється в відповідний медіатор, а потім транспортується в синаптичну щілину у вигляді бульбашок (везикул). Медіатори синтезуються так само в пресинаптичних закінченнях.
Механізм дії. Медіатори і модулятори зв'язуються з рецепторами постсинаптичної мембрани сусідніх клітин. Більшість нейромедіаторів стимулюють відкривання іонних каналів, і лише тільки деякі - закривання. Характер зміни мембранного потенціалу постсинаптичної клітини залежить від типу каналу. Зміна мембранного потенціалу від -60 до +30 мВ за рахунок відкривання Nа + -каналів призводить до виникнення постсинаптичного потенціалу дії. Зміна мембранного потенціалу з -60 мВ до -90 мВ за рахунок відкривання Cl - -каналів пригнічує потенціал дії (гиперполяризация), в результаті чого збудження не передається (гальмівний синапс). За хімічною будовою медіатори можна розділити на кілька груп, головними з яких є аміни, амінокіслоит, поліпептиди. Досить широко поширеним медіатором в синапсах ЦНС є ацетилхолін.
ацетилхолінзустрічається в різних відділах ЦНС (кора великого мозку, спинний мозок). Відомий в основному як збудливий медіатор. Зокрема, є медіатором альфа-мотонейронів спинного мозку, що іннервують скелетні м'язи. Цими нейронами передається збудливий вплив на гальмівні клітини Реншоу. У формації стовбура мозку, в гіпоталамусі виявлено М- і Н-холінорецептори. Ацетилхолін активує і гальмівні нейрони, що і визначає його ефект.
аміни (гістамін, дофамін, норадреналін, серотонін) в більшості своїй в значних кількостях міститися в нейронах стовбура мозку, в менших кількостях виявляються в інших відділах ЦНС. Аміни забезпечують виникнення процесів збудження і гальмування, наприклад, в проміжному мозку, чорної субстанції, лімбічної системи, смугастому тілі.
норадреналін. Норадренергические нейрони сконцентровані в основному в області блакитного плями (середній мозок), де їх налічується всього кілька сотень, але відгалуження їх аксонів зустрічаються по всій ЦНС. Норадреналін є гальмівним медіатором клітин Пуркіньє мозочка і збудливим - в гіпоталамусі, ядрах епіталямуса. У формації стовбура мозку і гіпоталамусі виявлено альфа і бета-адренорецептори. Норадреналін регулює настрій, емоційні реакції, забезпечує підтримку неспання, бере участь в механізмах формування деяких фаз сну, сновидінь.
Дофамін. Дофамінорецептори поділяють на Д1-і Д2- підтипи. Д1-рецептори локалізуються в клітинах смугастого тіла, діють за допомогою дофамінчувствітельной аденілатциклази, як і Д2-рецептори. Д2-рецептори виявлені в гіпофізі, при дії на них дофаміну пригнічуються синтез і секреція пролактину, окситоцину, меланостимулирующего гормону, ендорфіну . Дофамін бере участь у формуванні почуття задоволення, регуляції емоційних реакцій, підтримці неспання. Дофамін смугастого тіла регулює складні м'язові рухи.
Серотонін. За допомогою серотоніну в нейронах стовбура мозку передаються збуджуючі і гальмівні впливи, в корі мозку - гальмівні впливи. Є кілька типів серотонінових рецепторів. Серотонін реалізує свій вплив за допомогою іонотропних і метаботропних рецепторів, що впливають на біохімічні процеси за допомогою друге посередників - цАМФ і ІФ 3 / ДАГ. Міститься головним чином в структурах, що мають відношення до регуляції вегетативних функцій . Серотонін прискорює процеси навчання, формування больових відчуттів, сенсорне сприйняття, засипання; ангіотезінповишаетартеріальное тиск (АТ), гальмує синтез катехоламінів, стимулює секрецію гормонів; інформує ЦНС про осмотичний тиск крові.
гістамін в досить високій концентрації виявлено в гіпофізі і серединному підвищенні гіпоталамусу - саме тут сконцентровано основне кількість гістамінергіческіх нейронів. В інших відділах ЦНС рівень гістаміну дуже низький. Медіаторная роль його вивчена мало. Виділяють Н 1 -, Н 2 - і Н 3 гістамінорецепторов.
Амінокислоти.кислі амінокислоти (Гліцин, гамма-аміномасляна кислота) є гальмівними медіаторами в синапсах ЦНС і діють на відповідні рецептори. гліцин - в спинному мозку, ГАМК - в корі великого мозку, мозочку, стовбурі мозку і спинному мозку. нейтральні амінокислоти (Альфа- глутамат, альфа-аспартат) передають збуджуючі впливу і діють на відповідні збуджуючі рецептори. Припускають, що глутамат є медіатором аферентів в спинному мозку. Рецептори глутамінової та аспарагінової амінокислот є на клітинах спинного мозку, мозочка, таламуса, гіпокампа, кори великого мозку . Глутамат - основний збудливий медіатор ЦНС (75%). Рецептори глутамат іонотропние (K +, Ca 2+, Na +) і метаботропние (цАМФ і ІФ 3 / ДАГ). поліпептиди також виконують медіаторну функцію в синапсах ЦНС. Зокрема, субстанція Р є медіатором нейронів, що передають сигнали болю. Особливо багато цього полепіптіда міститься в дорсальних корінцях спинного мозку. Це дозволило припустити, що субстанція Р може бути медіатором чутливих нервових клітин в області їх перемикання на вставні нейрони.
Енкефаліни і ендорфіни - медіатори нейронів, які блокують больові імпульсацію. Вони реалізують свій вплив за допомогою відповідних опіатнихрецепторів, кіт особливо щільно розташовуються на клітинах лімбічної системи; багато їх так само на клітинах чорної субстанції, ядрах проміжного мозку і солетарного тракту, є вони на клітинах блакитного плями спинного мозга.Ендорфіни, енкефаліни, пептид викликає бета-сон, дають антибольовим реакції, підвищення стійкості до стресу, сон. ангіотензин бере участь у передачі інформації про потреби організму у воді, люліберін - в статевої активності. олігопептиди - медіатори настрою, статевої поведінки, передачі ноцицептивного збудження від периферії в ЦНС, формування больових відчуттів.
Хімічні речовини, які циркулюють в крові (Деякі гормони, простагландини, надають модулюючий вплив на активність синапсів. Простагландини (ненасичені оксикарбонові кислоти), що вивільняються з клітин, впливають на багато ланок синаптического процесу, наприклад на секрецію медіатора, роботу Аденилатциклаза. Вони володіють високою фізіологічною активністю, але швидко інактивуються і тому діють локально.
Гіпоталамічні нейрогормони,регулюють функцію гіпофіза, також виконують роль медіатора.
принцип Дейла. Згідно з цим принципом, кожен нейрон синтезує і використовує один і той же медіатор або одні і ті ж медіатори у всіх розгалуженнях свого аксона (один нейрон - один медіатор), але, як з'ясувалося, в закінченнях аксона можуть виділятися і інші, супутні, медіатори ( комедіатори), які відіграють модулирующую роль і діючі більш повільно. У спинному мозгеустановленно два швидкодіючих медіатора в одному гальмівному нейроне - ГАМК і гліцин, а також один гальмівний (ГАМК) і один збудливий (АТФ). Тому принцип Дейла в новій редакції звучить так: «один нейрон - один швидкий синаптический ефект». Ефект дії медіатора залежить в основному від властивостей іонних каналів постсинаптичної мембрани і друге посередників. Це явище особливо яскраво демонструється при порівнянні ефектів окремих медіаторів в ЦНС і периферичних синапсах організму. Ацетилхолін, наприклад, в корі мозку при мікроапплікаціях на різні нейрони може викликати збудження і гальмування, в синапсах серця - гальмування, в синапсах гладкої мускулатури шлунково-кишкового тракту - збудження. Катехоламіни стимулюють серцеву діяльність, але гальмують скорочення шлунка і кишечника.
З вищевикладеного зрозуміло, яке значення у функціях нервової системи відіграють медіатори. У відповідь на прихід нервового імпульсу до синапси відбувається викид медіатора; молекули медіатора з'єднуються (комплементарно - як «ключ до замка») з рецепторами постсинаптичної мембрани, що призводить до відкривання іонного каналу або до активування внутрішньоклітинних реакцій. Приклади синаптичної передачі, розглянуті вище, повністю відповідають цій схемі. Разом з тим завдяки дослідженням останніх десятиліть ця досить проста схема хімічної синаптичної передачі значно ускладнилася. Поява иммунохимических методів дозволило показати, що в одному синапсі можуть співіснувати кілька груп медіаторів, а не один, як це припускали раніше. Наприклад, в одному синаптическом закінчення одночасно можуть перебувати синаптичні пухирці, що містять ацетилхолін і норадреналін, які досить легко ідентифікуються на електронних фотографіях (ацетилхолін міститься в прозорих бульбашках діаметром близько 50 нм, а норадреналін - в електронно-щільних діаметром до 200 нм). Крім класичних медіаторів, в синаптическом закінчення можуть перебувати один або кілька ній-ропептідов. Кількість речовин, що містяться в синапсі, може доходити до 5-6 (своєрідний коктейль). Більш того, медіаторная специфічність синапсу може змінюватися в онтогенезі. Наприклад, нейрони симпатичних гангліїв, що іннервують потові залози у ссавців, початково норадренергічни, але у дорослих тварин стають холінергічних.
В даний час при класифікації медіаторних речовин прийнято виділяти: первинні медіатори, супутні медіатори, медіатори-модулятори і аллостерічеськіє медіатори. Первинними медіаторами вважають ті, які діють безпосередньо на рецептори постсинаптичної мембрани. Супутні медіатори і медіатори-модулятори можуть запускати каскад ферментативних реакцій, які, наприклад, фосфорилируют рецептор для первинного медіатора. Аллостерічеськіє медіатори можуть брати участь в кооперативних процесах взаємодії з рецепторами первинного медіатора.
Довгий час за зразок брали синаптичну передачу по анатомічній адресою (принцип «точка - в точку»). Відкриття останніх десятиліть, особливо медиаторной функції нейропептидів, показали, що в нервовій системі можливий принцип передачі і за хімічним адресою. Іншими словами, медіатор, що виділяється з даного закінчення, може діяти не тільки на «свою» постсинаптическую мембрану, а й за межами даного синапсу - на мембрани інших нейронів, що мають відповідні рецептори. Таким чином, фізіологічна реакція забезпечується не точним анатомічним контактом, а наявністю відповідного рецептора на клітці-мішені. Власне цей принцип був давно відомий в ендокринології, а дослідження останніх років знайшли йому більш широке застосування.
Всі відомі типи хеморецепторів на постсинаптичні мембрані поділяють на дві групи. В одну групу входять рецептори, до складу яких включено іонний канал, що відкривається при зв'язуванні молекул медіатора з «який дізнається» центром. Рецептори другої групи (метаботропние рецептори) відкривають іонний канал опосередковано (через ланцюжок біохімічних реакцій), зокрема, за допомогою активації спеціальних внутрішньоклітинних білків.
Одними з найпоширеніших є медіатори, що належать до групи біогенних амінів. Ця група медіаторів досить надійно ідентифікується мікрогістологіческімі методами. Відомі дві групи біогенних амінів: катехоламіни (дофамін, норадреналін і адреналін) і індоламін (серотонін). Функції біогенних амінів в організмі дуже різноманітні: медіаторная, гормональна, регуляція ембріогенезу.
Основним джерелом норадренергических аксонів є нейрони блакитного плями і прилеглих ділянок середнього мозку (рис. 2.14). Аксони цих нейронів широко поширюються в мозковому стовбурі, мозочку, в великих півкулях. У довгастому мозку велике скупчення норадренергических нейронів знаходиться в вентролатеральном ядрі ретикулярної формації. У проміжному мозку (гіпоталамусі) норадренергические нейрони поряд з дофаминергическими нейронами входять до складу гіпоталамо-гіпофізарної системи. Норадренергические нейрони в великій кількості містяться в периферичної нервової системи. Їх тіла лежать в симпатичної ланцюжку і в деяких інтрамуральних гангліях.
Дофамінергічні нейрони у ссавців знаходяться переважно в середньому мозку (так звана нігро-неостріарная система), а також в гіпоталамічної області. Дофамінові ланцюга мозку ссавців добре вивчені. Відомі три головні ланцюги, всі вони складаються з однонейронной ланцюжка. Тіла нейронів знаходяться в мозковому стовбурі і відсилають аксони в інші області головного мозку (рис. 2.15).

Одна ланцюг дуже проста. Тіло нейрона знаходиться в області гіпоталамуса і відсилає короткий аксон в гіпофіз. Цей шлях входить до складу гіпоталамо-гіпофізарної системи і контролює систему ендокринних залоз.
Друга дофаминовая система також добре вивчена. Це чорна субстанція, багато клітини якої містять дофамін. Аксони цих нейронів проектуються в смугасті тіла. Ця система містить приблизно 3/4 дофаміну головного мозку. Вона має вирішальне значення в регулюванні тонічних рухів. Дефіцит дофаміну в цій системі призводить до хвороби Паркінсона. Відомо, що при цьому захворюванні відбувається загибель нейронів чорної субстанції. Введення L-DOPA (попередника дофаміну) полегшує у хворих деякі симптоми захворювання.
Третя дофаминергическая система бере участь в прояві шизофренії та деяких інших психічних захворювань. Функції цієї системи поки вивчені недостатньо, хоча самі шляхи добре відомі. Тіла нейронів лежать в середньому мозку поряд з чорною субстанцією. Вони проектують аксони в вищерозміщені структури мозку, мозкову кору і лімбічну систему, особливо до фронтальній корі, до септальних області і енторінальной корі. Енторінальная кора, в свою чергу, є головним джерелом проекцій до гиппокампу.
Згідно дофаминовой гіпотезі шизофренії, третя дофаминергическая система при цьому захворюванні надактивного. Ці уявлення виникли після відкриття речовин, що знімають деякі симптоми захворювання. Наприклад, хлорпромазин і галоперидол мають різну хімічну природу, але вони однаково пригнічують активність дофаминергической системи мозку і прояв деякі симптомів шизофренії. У хворих на шизофренію, протягом року отримували ці препарати, з'являються рухові порушення, що отримали назву tardive dyskinesia (повторювані химерні руху лицьової мускулатури, включаючи мускулатуру рота, які хворий не може контролювати).
Серотонін майже одночасно відкрили в якості сироваткового сосудосуживающего фактора (1948) і ентераміна, що секретується ентерохромаффіновимі клітинами слизової оболонки кишечника. У 1951 р було розшифровано хімічну будову серотоніну і він отримав нову назву - 5-гідрокси. В організмі ссавців він утворюється гідроксилюванням амінокислоти триптофану з подальшим декарбоксилюванням. 90% серотоніну утворюється в організмі ентерохромаффіновимі клітинами слизової оболонки всього травного тракту. Внутрішньоклітинний серотонін інактивується моноаміноксидазою, що міститься в мітохондріях. Серотонін позаклітинного простору окислюється перулоплазміном. Велика частина виробленого серотоніну зв'язується з кров'яними пластинками і по кров'яному руслу розноситься по організму. Інша частина діє в якості місцевого гормону, сприяючи авторегулирования кишкової перистальтики, а також модулюючи епітеліальну секрецію і всмоктування в кишковому тракті.

Серотонінергічні нейрони широко поширені в центральній нервовій системі (рис. 2.16). Вони виявляються в складі дорсального і медіального ядер шва довгастого мозку, а також в середньому мозку і вароліевом мосту. Серотонінергічні нейрони іннервують великі ділянки мозку, що включають кору великих півкуль, гіпокамп, блідий шар, мигдалину, область гіпоталамуса. Інтерес до серотоніну був притягнутий в зв'язку з проблемою сну. При руйнуванні ядер шва тварини страждали безсонням. Подібний ефект надавали речовини, що виснажують сховище серотоніну в мозку.
Найвища концентрація серотоніну виявлена \u200b\u200bв епіфізі (pineal gland). Серотонін в епіфізі перетворюється в мелатонін, який бере участь в пігментації шкіри, а також впливає у багатьох тварин на активність жіночих гонад. Зміст як серотоніну, так і мелатоніну в епіфізі контролюється циклом світло - темрява через нервову симпатичну систему.
Іншу групу медіаторів ЦНС складають амінокислоти. Вже давно відомо, що нервова тканина з її високим рівнем метаболізму містить значні концентрації цілого набору амінокислот (перераховані в порядку убування): глутамінової кислоти, глутаміну, аспарагінової кислоти, гамма-аміномасляної кислоти (ГАМК).
Глутамат в нервовій тканині утворюється переважно з глюкози. У ссавців найбільше глутамату міститься в кінцевому мозку і мозочку, де його концентрація приблизно в 2 рази вище, ніж в стовбурі мозку і спинному мозку. У спинному мозку глутамат розподілено нерівномірно: в задніх рогах він знаходиться в більшій концентрації, ніж в передніх. Глутамат є одним з найпоширеніших медіаторів в ЦНС.
Постсинаптические рецептори до глутамату класифікуються відповідно до аффинностью (спорідненістю) до трьох екзогенних агонистам - квісгулату, каїнатом і N-метил-D-аспартату (NMDA). Іонні канали, що активуються квісгулатом і каїнатом, подібні каналам, які управляються нікотиновими рецепторами - вони пропускають суміш катіонів (Na + і. До +). Стимуляція NMDA-рецепторів має складний характер активації: іонний струм, який переноситься не тільки Na + і К +, але також Са ++ при відкриванні іонного каналу рецептора, залежить від потенціалу мембрани. Вольтзавісімая природа цього каналу визначається різним ступенем його блокування іонами Mg ++ з урахуванням рівня мембранного потенціалу. При потенціалі спокою порядку - 75 мВ іони Mg ++, які переважно знаходяться в міжклітинному середовищі, конкурують з іонами Са ++ і Na + за відповідні канали мембрани (рис. 2.17). Внаслідок того, що іон Mg ++ не може пройти через пору, канал блокується щоразу, як потрапляє туди іон Mg ++. Це призводить до зменшення часу відкритого каналу і провідності мембрани. Якщо мембрану нейрона деполярізовать, то кількість іонів Mg ++, які закривають іонний канал, знижується і через канал безперешкодно можуть проходити іони Са ++, Na + і. До +. При рідкісних стимуляції (потенціал спокою змінюється мало) глутаматергіческіх рецептораВПСП виникає переважно за рахунок активації квісгулатних і каінатного рецепторів; внесок NMDA-рецепторів незначний. При тривалій деполяризації мембрани (ритмічна стимуляція) магнієвий блок видаляється, і NMDA-канали починають проводити іони Са ++, Na + і. До +. Іони Са ++ через вторинні посередники можуть потенціювати (посилювати) мінПСП, що може привести, наприклад, до тривалого збільшення синаптичної провідності, що зберігається годинами і навіть цілодобово.

З гальмівних медіаторів ГАМК є найпоширенішою в ЦНС. Вона синтезується з L-глутамінової кислоти в одну стадію ферментом декарбоксилаз, наявність якої є лімітуючим фактором цього медіатора. Відомо два типи ГАМК-рецепторів на постсинаптичні мембрані: ГАМКА (відкриває канали для іонів хлору) і ГАМКб (відкриває в залежності від типу клітини канали для. До + або Са ++). На рис. 2.18 показана схема ГАМК-рецептора. Цікаво, що в його склад входить бензодіазіпіновий рецептор, наявністю якого пояснюють дію так званих малих (денних) транквілізаторів (седуксен, тазепама і ін.). Припинення дії медіатора в ГАМК-синапсах відбувається за принципом зворотного всмоктування (молекули медіатора спеціальним механізмом поглинаються з синаптичної щілини в цитоплазму нейрона). З антагоністів ГАМК добре відомий бікукулін. Він добре проходить через гематоенцефалічний бар'єр, робить сильний вплив на організм навіть в малих дозах, викликаючи конвульсії і смерть. ГАМК виявляється в ряді нейронів мозочка (в клітинах Пуркіньє, клітинах Гольджі, корзинчатих клітинах), гіпокампу (в корзинчатих клітинах), в нюхової цибулини і чорної субстанції.
Ідентифікація ГАМК-ланцюгів мозку важка, так як ГАМК - звичайний учасник метаболізму в ряді тканин організму. Метаболічна ГАМК не використовується як медіатор, хоча в хімічному відношенні їх молекули однакові. ГАМК визначається по ферменту декарбоксилази. Метод заснований на отриманні у тварин антитіл до декарбоксилази (антитіла екстрагують, мітять і вводять в мозок, де вони зв'язуються з декарбоксилаз).

Іншим відомим гальмівним медіатором є гліцин. Гліцинергічними нейрони знаходяться головним чином в спинному і довгастому мозку. Вважають, що ці клітини виконують роль гальмівних интернейронов.

Ацетилхолін - один з перших вивчених медіаторів. Він надзвичайно широко поширений в периферичної нервової системи. Прикладом можуть служити мотонейрони спинного мозку і нейрони ядер черепних нервів. Як правило, холинергические ланцюга в мозку визначають по присутності ферменту холінестерази. У головному мозку тіла холинергических нейронів знаходяться в ядрі перегородки, ядрі діагонального пучка (Брока) і базальних ядрах. Нейроанатомии вважають, що ці групи нейронів формують фактично одну популяцію холинергических нейронів: ядро \u200b\u200bпеднего мозку, nucleus basalis (воно розташоване в базальної частини переднього мозку) (рис. 2.19). Аксони відповідних нейронів проектуються до структур переднього мозку, особливо в нову кору і гіпокамп. Тут зустрічаються обидва типи ацетилхолінових рецепторів (мускаринові і нікотинові), хоча вважається, що мускаринові рецептори домінують в більш рострально распоженних мозкових структурах. За даними останніх років складається враження, що ацетилхолінового система відіграє велику роль в процесах, пов'язаних з вищими інтегративними функціями, які вимагають участі пам'яті. Наприклад, показано, що в мозку хворих, які померли від хвороби Альцгеймера, спостерігається масивна втрата холінергічних нейронів в nucleus basalis.
ГАМК - гамма-аміномасляна кислота - є головним гальмівним нейромедіатором в головному мозку, вона причетна як до постсинаптичного, так і до пресинаптичного гальмування. ГАМК утворюється з глутамату під впливом глутаматдекарбоксилази і взаємодіє з двома типами ГАМК-рецепторів постсинаптичних мембран синапсів: а) при взаємодії з ГАМКд-рецепторами збільшується проникність іонних каналів мембрани для іонів СГ, що має місце в клінічній практиці при застосуванні барбітуратів; б) при взаємодії з ГАМК-рецепторами збільшується проникність іонних каналів для іонів К +. гліцин - гальмівний нейромедіатор, виділяється переважно нейронами спинного мозку і стовбура мозку. Він збільшує провідність іонних каналів постсинаптичної мембрани для іонів СГ, що призводить до розвитку гіперполяризації - ГПСП. Антагоністом гліцину є стрихнін, введення якого призводить до м'язової гіперактивності і судом, що підтверджує важливу роль постсинаптичного гальмування в нормальної функції центральної нервової системи. Правцевий токсин теж призводить до виникнення судом. Діючи на білок сінаптобревін мембран везикул, він блокує екзоцитоз пресинаптического гальмівного нейромедіатора, в результаті чого виникає різке збудження ЦНС.
Міжнейронних передача збудження може відбуватися також електричним шляхом, тобто без участі медіаторів. Умовою для цього є щільний контакт між двома клітинами шириною до 9 нм. Отже, натрієвий струм від однієї з них може проходити через відкриті канали інший мембрани. Тобто джерелом постсинаптичного струму другого нейрона є пресинаптическая мембрана першого. Процес безмедіаторній; забезпечується виключно канальними білками (ліпідні мембрани для іонів непроникні). Саме такі міжклітинні зв'язку названий Нексус (щілинними контактами). Вони розташовані строго один проти одного в мембранах двох нейронів - тобто на одній лінії; діаметром великі (до 1,5 нм в поперечнику), пропускають навіть для макромолекул масою до 1000 Складаються з субодиниць масою до 25000, їх наявність звичайна для ЦНС як хребетних, так і безхребетних; властива групам синхронно функціонуючих клітин (зокрема, знайдені в мозочку між клітинами-зернами).
Більшість електричних синапсів є збудливими. Але при певних морфологічних характеристиках вони можуть бути гальмівними. При двобічності проведення деякі з них мають випрямний ефект, тобто проводять електричний струм значно краще пресинаптических структур до постсинаптических, ніж у зворотному напрямку.
Кожен нервовий центр має свою морфологічну і функціональну специфіку. Але в основі нейродинамики будь-якого з них закладено ряд спільних рис. Вони пов'язані з механізмами передачі збудження в синапсах; з взаємодією між нейронами, які входять до складу цього центру; з генетично запрограмованими функціональними особливостями нейронів і зв'язків між ними.
Особливості проведення збудження через синапси такі.
1 Однобічність проведення збудження. У аксоні збудження проходить в обох напрямках від місця його виникнення, в нервовому центрі - тільки в одному напрямку: від рецептора до ефектора (тобто на рівні синапсу від пресинаптичної мембрани до постсинаптичні), що пояснюється структурно-функціональної організацією синапсу, а саме - відсутністю синаптичних пухирців з медіатором в постсинаптичних нейронах, 2 Сннаптнчна затримка проведення збудження. збудження в нервовому центрі проводиться з меншою швидкістю, ніж в інших частинах рефлекторної дуги. Це пов'язано з тим, що витрачається на процеси виділення медіатора, з фізико-хімічними процесами, які відбуваються в синапсі, з виникненням ВПСП і генерацією ПД. На все це в одному синапсі витрачається 0,5-1 мс. Таке явище отримало назву синаптичної затримки проведення збудження. Чим складніше рефлекторна дуга, тим більше синапсів і, відповідно, більша синаптическая затримка.
Сума синаптичних затримок в рефлекторну дугу отримала назву теперішнього часу рефлексу. Час від початку дії подразника до появи рефлекторного відповіді називається прихованим, або латентним періодом (ЛП) рефлексу. Тривалість цього періоду залежить від кількості нейронів, а отже і синапсів, що беруть участь в рефлексі. Наприклад, сухожильний колінний рефлекс, рефлекторна дуга якого моносинаптичних, має ЛП 24 мс, зорова або слухова реакція - 200 мс.
Залежно від того, що збуджують або гальмівні нейрони здійснюють синаптичні контакти, сигнал може посилюватися або придушуватися. Механізми взаємодії між збудливими і гальмівними впливами на нейроні лежать в основі їх інтегративної функції.
Таким механізмом взаємодії є підсумовування на нейроні збуджуючих впливів - збудливий постсинаптичний потенціал (ВПСП), або гальмівних впливів - гальмівний постсинаптичний потенціал (ТПСП), або одночасно збуджуючих (ВПСП) і гальмівних (ГПРП).
3 Суммация нервових процесів - явище виникнення збудження за певних умов нанесення підпорогових подразнень. Підсумовування описано І. М. Сеченова. Виділяють два види сумації: тимчасова сумація і просторова сумація, (рис. 3.15).
часова сумація - виникнення збудження на ряд підпорогових подразнень, послідовно надходять в клітину або центру від одного рецепторного поля (рис. 3.16). Частота стимулів повинна бути такою,
МАЛ. 3.15. Суммация збудження. А - тимчасова сумація. Б - просторова сумація

МАЛ. 3.16.
щоб інтервал між ними був не більше 15 мс, тобто коротше тривалість ВПСП. При таких умовах ВПСП на черговий стимул розвивається до того, як завершиться ВПСП на попередній стимул. ВПСП підсумовуються, їх амплітуда зростає і, нарешті, при досягненні критичного рівня деполяризації, виникає ПД.
просторова сумація - виникнення збудження (ВПСП) при одночасному нанесенні декількох допорогових стимулів на різні ділянки рецепторного ПОЛЯ (рис. 3.17).
Якщо ВПСП виникають одночасно в кількох синапсах нейрона (не менше 50), мембрана нейрона деполяризуется до критичних значень і, як наслідок, виникає ПД. Просторова сумація процесів збудження (ВПСП) і гальмування (ГПРП) забезпечує інтегративну функцію нейронів. Якщо переважає гальмування, інформація не передається до наступного нейрона; якщо переважає збудження - інформація передається далі до наступного нейрона завдяки генерації ПД на мембрані аксона (рис. 3.18).
4 Трансформація ритму збудження - це невідповідність частоти ПД в аферентних і еферентних ланках рефлекторної дуги. Наприклад, у відповідь на одиночний стимул, прикладений

МАЛ. 3.17.

МАЛ. 3.18.
до аферентного нерва, центри по еферентних волокнах посилають до робочого органу цілу серію імпульсів один за іншим. В іншій ситуації при великій частоті стимуляції до ефектора надходить значно менше частота.
5 Післядія збудження - явище продовження порушення в ЦНС після припинення подразнення. Короткочасна післядія пов'язана з великою тривалістю ВПСП критичного рівня. Тривала післядія обумовлена \u200b\u200bциркуляцією збудження замкнутими нервовими ланцюгами. Таке явище називається реверберацией. Завдяки реверберації збуджень (ПД) нервові центри постійно знаходяться в стані тонусу. Розвиток реверберації на рівні цілісного організму важливий при організації пам'яті.
6 Постгетанічна потенциация - явище появи або посилення відповіді на окремі тестуючі сенсорні стимули протягом деякого часу після попереднього слабкого частого (100-200 НМЛ / с) ритмічного подразнення. Потенціація обумовлена \u200b\u200bпроцесами а рівні пресинаптичної мембрани і виражається збільшенням виділення медіатора. Це явище має гомосінаптічну природу, тобто виникає в тому випадку, коли ритмічне роздратування і тестує імпульс надходять до нейрона по одним і тим же аферентних волокнах. В основу потенциации належить, перш за все, зростання надходження Ca2f через пресинаптичної мембрани. Це явище прогресивно зростає з кожним імпульсом. І коли кількість Са 2+ стає більше, ніж здатність мітохондрій і ендоплазматичної їх абсорбувати, настає пролонговану звільнення медіатора в синапс. Отже, має місце мобілізація готовності до виділення медіатора великою кількістю везикул і як наслідок - збільшення кількості квантів медіатора на постсинаптичні мембрані. За сучасними даними, в генезі посттетанічного потенциации велику роль відіграє секреція ендогенних нейропептидів, особливо при переході короткочасної потенциации в довготривалу. Серед них знаходяться нейромодулятора, що діють як на пресинаптическую, так і на постсинаптичні мембрані. Стимуляторами є соматостатин, фактор росту, а інгібіторами - інтерлейкін, тіроліберін, мелатонін. Значні також арахідонової кислоти, NO. Потенціація має значення при організації пам'яті. Завдяки підсилює ланцюгах організовується навчання.
7 стомлюваність нервових центрів. При тривалому повторному виконанні того ж рефлексу через деякий час настає стан зменшення сили рефлекторної реакції і навіть повне її придушення, тобто настає втома. Втома в першу чергу розвивається в нервовому центрі. Вона пов'язана з порушенням передачі в синапсах, виснаженням ресурсів медіатора в пресинаптичних везикулах, зниженням чутливості рецепторів субсінаптічноі мембрани до медіаторів, а також ослабленням дії ферментних систем. Однією з причин є "" звикання "" постсинаптичної мембрани до дії медіатора - габітуація.
Деякі хімічні речовини специфічно впливають на відповідні нервові центри, що пов'язано зі структурами цих хімічних речовин, які можуть бути спорідненими з відповідними медіаторами нервових центрів.
Серед них:
1 наркотізуючі - такі, що використовуються в хірургічній практиці для наркозу (хлоретил, кетамін, барбітурати та ін.);
2 транквілізатори - заспокійливі засоби (реланіум, аміназин, триоксазин, амизил, оксілідін, серед рослинних препаратів - настій пустирника, півонії та ін.);
3 нейротропні речовини вибіркової дії (лобелін, цититон - збудники дихального центру; апоморфин - збудник центру блювоти; мескалін - зоровий галюциноген і ін.).
медіатори (Трансмітери) - фізіологічно активні речовини, безпосередньо передають інформацію від однієї клітини до іншої через спеціальні міжклітинні контакти - синапси.
На периферії медіаторами найчастіше служать два речовини - АХ (нервово-м'язові синапси і синапси парасимпатичного відділу ВНС) і НА (синапси постгангліоіарних волокон симпатичного відділу ВНС). Але в ЦНС збудження і гальмування можуть передаватися з нейрона на нейрон за допомогою багатьох медіаторів. Серед збуджуючих медіаторів найчастіше зустрічаються глутамат, АХ, НА, Д, серотонін, а серед гальмівних - ГАМК і гліцин. Але є і досить рідко зустрічаються хімічні посередники, що виробляються в відносно малому числі нервових клітин. Вважають, що медіаторами в нашому мозку є не менше 35-40 різних речовин. Саме порушення у виробленні або утилізації медіаторів є основною причиною безлічі нервових і психічних розладів.
Властивості речовини, здатного стати медіатором, представлені на рис. 9.4.
Мал. 9.4.
1 - медіатор і його хімічні попередники повинні бути присутніми в нейроні; 2 - медіатор повинен міститися в високих концентраціях в синаптичних везикулах; 3 - в синаптическом закінчення і (або) в тілі нейрона повинна міститися ферментативна система синтезу медіатора; 4 - медіатор повинен вивільнятися з везикул в синаптичну щілину при приході ПД до нервового закінчення; 5 - викиду медіатора в синаптичну щілину при стимуляції повинен передувати вхід в закінчення іонів кальцію; 6 - в синаптичної щілини повинна бути присутнім система деградації медіатора і (або) система його зворотного захоплення в пресинаптичне закінчення; 7 - на постсинаптичні мембрані повинні бути присутніми рецептори до медіатора
за своєю хімічною природою медіатори можна поділити на « класичні», Що є видозміненими амінокислотами, і« некласичні»- пептидні і газоподібні (табл. 9.1). Традиційно медіатори ІА і Д, синтезовані в організмі з харчової амінокислоти фенілаланіну, що містить катехоловое ядро, називають катехоламинами. Серотонін, який синтезується з амінокислоти триптофану і за своєю хімічною природою представляє собою похідне індолу, разом з НА та Д відносять до групи біогенних амінів, хоча і серед інших медіаторів зустрічається безліч «амінів».
Таблиця 9.1
Деякі медіатори, виявлені в організмі тварин
За своїм ефектам класичні медіатори поділяють на збуджуючі і гальмівні. Багато пізніше «класичних» медіаторів були відкриті пептидні медіатори, що представляють собою невеликі ланцюжки з амінокислот. Доведено медіаторная роль кількох пептидів, і кілька десятків пептидів знаходяться «під підозрою». І нарешті, досить несподіваним було виявлення здатності клітин виробляти ряд газоподібних речовин, для секреції яких не потрібні «упаковка» в везикули; проте вони є повноцінними медіаторами. Краще за інших газів в ролі медіатора відомий оксид азоту (NO), але медіаторні властивості СО і H 2 S також не викликають сумнівів.
Будь-медіатор, незалежно від хімічної або фізичної природи, має свій життєвий цикл, Що включає наступні етапи:
Синтез медіаторів може відбуватися як в тілі нейрона, так і в самих пресинаптических закінченнях. Молекули медіаторів пептидної природи ферментативно «вирізаються» з великих білків-попередників, які синтезуються в тілі нейрона на шорсткою ЕПР. потім ці
медіатори упаковуються в апараті Гольджі в великі везикули, які за допомогою аксонального транспорту рухаються але аксону до синапсах. «Класичні» медіатори синтезуються в самому кінці його, куди ферменти для синтезу і упаковки молекул в везикули надходять за рахунок аксонального транспорту. У більшості нейронів домінує якийсь один медіатор, проте в останні роки встановлено, що в одному і тому ж нейроні і, більш того, в одному і тому ж синапсі можуть бути присутні кілька медіаторів. Вони можуть перебувати як в одних і тих же, так і в різних везикулах. Таке співіснування показано, наприклад, для біогенних амінів і пептидних медіаторів.
Викид медіатора в синаптичну щілину відбувається в той момент, коли ПД досягає нервової терміналі і пресинаптическая мембрана деполяризуется (рис. 9.5).

Мал. 9.5.
У цей момент в мембрані відкриваються потенціалзалежні кальцієві канали і Са 2+ входить в пресинаптичне закінчення, зв'язуючись з певним білком на зовнішній стороні мембрани везикул і запускаючи процес злиття везикули і пресинаптичної мембрани. Везикула може, по-перше, вбудуватися в неї цілком і «викинути» весь свій вміст в синаптичну щілину ( «повне злиття»). По-друге, може утворюватися короткостроковий контакт ( «пора злиття») зі спеціальних білків між мембраною везикули і мембраною закінчення. Через пору злиття частина молекул медіатора встигає вийти в синаптичну щілину (такий спосіб секреції медіатора називається « kiss-and-run»(В перекладі з англ,« цілуй і біжи »).
Як тільки медіатор виявився в щілини, необхідно дуже швидко прибрати увійшов в нервове закінчення кальцій. Для цього існують спеціальні зв'язують кальцій білки-буфери, а також кальцієві насоси, що відкачують кальцій в ендоплазматичну мережу, в мітохондрії і в зовнішнє середовище. В цей час спустошені ( kiss-and-run) Або заново формуються в нервовому закінченні везикули знову наповнюються молекулами медіатора.
Потрапили в синаптичну щілину молекули медіатора досягають постсинаптичної мембрани за рахунок дифузії і взаємодіють з рецепторами. Традиційно терміном «рецептор» позначаються спеціальні клітини або клітинні чутливі освіти, що реагують на подразники зовнішнього і внутрішнього середовища: фоторецептори, механорецептори і т.п. У сучасній біології термін «рецептор» вживають також по відношенню до молекул білків, вбудованих в клітинну мембрану або знаходяться в цитоплазмі і здатних реагувати зміною своєї форми і стану па специфічні для кожного виду рецепторів впливу. Рецептори знайдені для медіаторів, гормонів, антитіл та інших сигнальних молекул, важливих для передачі інформації в живих системах.
Передача сигналу через мембрану включає в себе три стадії:
виділяють два основних види рецепторів - іонотропние (канальні) і метаботропние.
прикладом канального рецептора може служити ліганд-активується (хемочувствітельності) рецептор для АХ, що розташовується на мембрані волокон скелетних м'язів (див. рис. 8.17). Такі рецептори крім природного АХ активуються алкалоїдом тютюну - нікотином. Тому їх називають нікотиновими або Н-холинорецепторами. Крім поперечно-смугастих м'язів такі рецептори зустрічаються і в ЦНС. Канал складається з п'яти білкових субодиниць, зібраних в своєрідну трубку, пронизливий мембрану наскрізь. Дві субодиниці однакові і позначаються а. Коли дві молекули медіатора АХ приєднуються до спеціальних місцях зв'язування на а-субодиниць, канал відкривається для катіонів Na + і Са 2+ (рис. 9.6).
В результаті на постсинаптичні мембрані розвивається ВПСП, і клітина може збудитися. Взаємодія медіатора з рецептором триває 1-2 мс, а потім молекула медіатора повинна від'єднатися, інакше рецептор «втратить чутливість» і тимчасово перестане реа-
гірованного на нові порції медіатора. Канальний вид рецепції дуже швидкий, проте він зводиться або до деполяризації постсинаптичної клітини за допомогою відкриття катіонних каналів, або до се гіперполяризації шляхом відкриття хлорних каналів.

Мал. 9.6.
а - схема будови; 6 - капа.;: Закритий; в - канал відкритий; А - ангстрем (1СГ 10 м)
метаботропние рецептори являють собою молекули білків, які сім разів «пропущені» через клітинну мембрану, утворюючи три петлі всередині клітини і три - на зовнішній стороні клітинної мембрани (рис. 9.7).

Мал. 9.7.
а, р, у - субодиниці G-бел ка
В даний час виявлено безліч подібних рецепторних білків, причому частина білкової молекули, звернена всередину клітини, пов'язана з відповідним G-білком. G-білки отримали свою назву за здатність розщеплювати ГТФ (гуанозинтрифосфат) до ГДФ (гуанозиндифосфат) і залишку фосфорної кислоти. Ці білки складаються з трьох субодиниць: а, р, у (див. Рис. 9.7), причому відомо кілька підтипів а-субодиниць. Той чи інший підтип а-субодиниць, що входять до складу G-білка, визначає, на який процес в клітці буде впливати даний G-білок. Наприклад, Gj.-білок (тобто включає в себе а 5 -субодиниці) стимулює фермент АЦ, G q стимулює фосфоліпазу С, G 0 зв'язується з іонними каналами, Gj гальмує активність АТ. Часто один різновид G-білка впливає на кілька процесів в клітині. За відсутності ліганду (медіатора або гормону), який може зв'язатися з метаботропних рецептором, G-білок неактивний. Якщо ж з рецептором зв'язався відповідний йому активує ліганд - а-субодиниця активується (ГДФ заміщується на ГТФ), від'єднується від комплексу субодиниць Ру і короткий час взаємодіє з білками-мішенями, запускаючи або, навпаки, гальмуючи внутрішньоклітинні процеси. Субодиниці G-білка не можуть існувати окремо протягом довгого часу і після гідролізу ГТФ а-суб'едіііцей утворюють єдиний неактивний G-білок. Діючи на цілий ряд ферментів і іонних каналів, активовані G-білки запускають каскад внутрішньоклітинних хімічних реакцій, в результаті яких змінюється концентрація низки регуляторних молекул - вторинних посередників (Первинні посередники - молекули, які переносять сигнал від клітини до клітини, Г.Є. медіатор, гормон).
До найбільш поширених вторинним посередникам (мессенджерам) відносять цАМФ, який утворюється з АТФ під дією ферменту АЦ. Якщо ж в результаті впливу ліганда на рецептор активується G ^-форма білка, то вона активує фермент фосфоліпазу С, яка в свою чергу стимулює утворення з фосфоліпідів мембран двох посередників: ІФ 3 (інозітолтріфосфат) і ДАГ (діацілгліцерола). Обидва посередника призводять до збільшення в клітці концентрації кальцію за рахунок його надходження ззовні (через іонні канали) або при викиді його з внутрішньоклітинних депо. Са 2+ - найпотужніший внутрішньоклітинний стимулятор процесів життєдіяльності клітини. Крім того, ІФ 3 і ДАГ стимулюють ріст клітин, сприяють експресії генів, вивільненню медіаторів, секреції гормонів і т.п. Однак вторинний посередник прямо або через ряд проміжних стадій впливає на хемочувствітельності іонні канали - відкриває або закриває їх. Це сприяє розвитку збудження або гальмування клітини, в залежності від того, які канали зазнали впливу. Величина і тривалість потенціалів буде залежати від виду, кількості та часу взаємодії молекул медіатора з рецепторами, а в підсумку - від того, яка система вторинних посередників активується під дією медіатора.
Характерною рисою метаботропних рецепції є її каскадність, що дозволяє багаторазово посилити вплив медіатора на клітку (рис. 9.8).

Мал. 9.8.
Як вже говорилося, медіатор не повинен взаємодіяти з іонотропних або метаботропних рецептором довше 1-2 мс. У нервово-м'язових синапсах АХ дуже швидко руйнується ферментом ацетилхолінестеразою до холіну і ацетату. Утворився холін транспортується в пресінап- тическое закінчення і знову використовується для синтезу АХ. Аналогічно руйнуються відповідними ферментами в синаптичної щілини і інші медіатори (АТФ, пептиди).
Інший поширений варіант усунення медіатора з синаптичної щілини - це його зворотне захоплення (англ, reuptake) В пресинаптичне закінчення або в гліальні клітини. НА, Д і серотонін після захоплення закінченнями знову «пакуються» в везикули або можуть бути зруйновані внутрішньоклітинними ферментами. ГАМК і глутамат транспортуються з синаптичної щілини в клітини глії і, зазнавши ряд біохімічних перетворень, знову потрапляють в нервові закінчення.
В процесі еволюції природа створила безліч фізіологічно активних речовин, що діють на метаболізм медіаторів. Багато таких речовин виробляють рослини з метою захисту. Разом з тим, отрути, що діють на життєвий цикл медіаторів і синаптичну передачу, виробляють деякі тварини: для нападу на жертву або для оборони від хижаків.
Величезна кількість хімічних сполук, що впливають на роботу медіаторних систем, створюється штучно людиною в пошуках нових лікарських засобів, що впливають на функціонування НС.
| Статті по темі: | |
|
Гідридна система зберігання водню Відмінності фенолів від спиртів
Болл і пряник діаграма заліза (I) Гідриди (FeH) вільна молекула .... Кафедра колоїдної хімії імені С
зав. кафедройДоктор хімічних наук, старший науковий співробітник ... Характерні хімічні властивості вуглеводнів
Утворюється при перекривання атомних орбіталей і утворення загальних ... | |