Čitatelia Vibir




Populárna štatistika
Fotosyntéza - cena procesov pri syntéze organických z neekologických rastlín, premena svetelnej energie na energiu chemických spojení. Pred fototrofnými organizmami by mali byť zelené pruhy, deyakі prokarіoti - cyanobaktérie, fialové a zelené sírové baktérie, roslinnі dzhgutikovі.
Pred procesom fotosyntézy sa otriasli v druhej polovici XVIII. Ešte dôležitejšie je, že ruské učenie vidkritya zrobiv vidatny K.A. Ospalá glazúra Roslini vymieňa a premieňa svetelnú energiu na energiu chemických zvukov, ktoré syntetizujú v organických spolucách. Sám Tim vezme ten smrad, aby zachoval vývoj života na Zemi. Výskum tiež teoreticky ohraničil a experimentálne dokázal úlohu chlorofylu v íle v procese fotosyntézy.
Chlorofyly sú hlavné vo fotosyntetických chemikáliách. Pre štruktúru smradu je hemoglobín upravený na hemoglobín; Namiesto potrebnej prípravy na syntézu molekúl chlorofylu. Existuje malé množstvo chlorofylov, ktoré rastú vlastnou chémiou. Vyžaduje sa pre všetky fototrofie є chlorofyl a . Chlorofylb sedieť pri zelenom roseline, chlorofyl s - v rozsievkach a norách rias. Chlorofyl d charakteristické pre červené riasy.
Zelené a fialové fotosyntetické baktérie môžu byť špeciálne bakteriochlorofyl ... Fotosyntéza baktérie malígnej miechy z fotosyntézy roslínu. Výsledkom je, že darca baktérií má vodu a vodu a ruženín má vodu. Zelené a fialové baktérie nemajú fotosystém II. Bakteriálna fotosyntéza nie je kontrolovaná vizuálnymi obrazmi. Zhrnutie bakteriálnej fotosyntézy:
6C02 + 12H2S -> C6H1206 + 12S + 6H20.
Základom fotosyntézy je oxidačný proces. Od prevodu elektrónov od prevodu elektronického darcu od dodávateľa elektronického darcu do dňa prevodu, od prevodu akceptora. Energia Svitlovej sa premieňa na energiu syntetizovaného organického spoluku (v sacharidoch).
Na chloroplastových membránach є špeciálne štruktúry - reakčné centrá , scho revenge chlorofyl. Nech zelené vrčí a sinice rastú dvi fotosystémy – perchu (ja) і priateľ (II) , pretože môže dôjsť k zmene reakcie centra a prepojené medzi sebou prostredníctvom elektronického prenosového systému.
Proces fotosyntézy prebieha v dvoch fázach: svetlo a tma.

Byť zbavený prítomnosti svetla na vnútorných membránach mitochondrií v membránach špeciálnych štruktúr - tylakoidiv ... Fotosyntetické pigmenty zachytávajú kvantové svetlá (fotóny). Tse produkovať až "zabudzhennya" jeden z elektrónov molekuly chlorofylu. Pomocou molekúl-nosičov sa elektrón presunie na posledný povrch membrány tylakoidov, čím sa nafúkne spevácka potenciálna energia.
Tsey elektron u fotosystémy I Môžete zapnúť svoj vlastný energetický rіven a zmeniť názor. Môže sa prenášať aj NADP (nikotínadadeninukleotid fosfát). Vzaєmodiyuchi s іons vodnyu, elektroni vіdnovlyuyut tsyu spoluku. Obnova NADP (NADP H) je dodanie obnovy atmosférického CO2 glukóze.
Ďalšie procesy sa vykonávajú v fotosystém II ... Napájanie elektroniky môže byť prenášané fotosystémom a aktualizované. Pre predajňu elektroniky bude použitá renovácia fotosystému ІІ, ktorý bude zásobovať molekuly vody. Molekuly vody a štiepenie (fotoliz vodi) na protóne je molekulárna kyselina, ktorú možno vidieť v atmosfére. Electroni vikoristovuyt za aktualizáciu fotosystému II. Rivnyannya photolizu vodi:
2H20 -> 4H++ 02 + 2e.
Keď je elektronika otočená z vonkajšieho povrchu tylakoidnej membrány, energia je viditeľná na úrovni prednej energie. Musíte uložiť chemické väzby molekúl ATP, ktoré sa každú hodinu syntetizujú reakciami oboch fotosystémov. Proces syntézy ATP z ADP a kyseliny fosforečnej sa nazýva fotofosforylácia ... Deyaka je súčasťou energie vikorystovutsya pre viparovannya vodi.
Počas svetelnej fázy fotosyntézy je veľa energie na energiu: ATP a NADP H. Keď molekuly padajú (fotolýza), molekulárne lastúrniky sú viditeľné v atmosfére.

Reakcie prebiehajú proti vnútornému prostrediu chloroplastov. Môžete vidieť jaka pre prítomnosť svetla a bez neho. Organická reč sa syntetizuje (C0 2 sa obnovuje na glukózu) zo zdrojov energie, ktorá vznikla vo fáze svetla.
Proces obnovy v oxide uhličitom je cyklický a tzv Calvinov cyklus ... Pomenovaný na počesť amerického pastora M. Kalvina, čo je akýsi cyklický proces.
Na opravu cyklu reakcie atmosférického oxidu uhličitého s ribulózou bifosfátom. Enzým katalyzujúci proces karboxyláza ... Ribulóza bifosfát je reťazec p'iativcarbon zukor, s dvoma prebytkami kyseliny fosforečnej. Pridáva sa rad nízkych chemických konverzií, kožou katalyzované špecifické enzýmy. Yak kintseviy produkt fotosyntézy je akceptovaný glukózy ako aj zavedenie ribulózafosfátu
Sumarne k procesu fotosyntézy:
6C02 + 6H20 → C6H12Pro6 + 602

Manažéri procesu fotosyntézy sú glazovaní na svetelnú energiu Sontsya a v procese rekonfigurácie chemických spojení v syntetizovaných sacharidoch. Prostredníctvom lancerov sa energia prenáša na heterotrofné organizmy. V procese fotosyntézy sa naň viaže plynný oxid uhličitý a je vidieť bozk. Všetky atmosférické bozky majú fotosyntetickú podobnosť. Je bežné vidieť viac ako 200 miliárd ton vína. Kisen dobytie života na Zemi z ultrafialového viprominuvannya, blikajúca ozónová obrazovka atmosféry.
Proces fotosyntézy je neúčinný, fragmenty v syntetizovanej organickej reči sa prenášajú bez 1-2% ospalej energie. Je zviazaný tse z tim, no, výrastky nestačia na zasklenie svetla, ich časť, ktorú má zasklievať atmosféra, je príliš tenká. Väčšina ospalého svetla sa prebúdza zo Zeme späť do vesmíru.
1. Ujasnite si hodnotu.
Fotosyntéza- proces vytvárania organickej reči z oxidu uhličitého a rozsvietenia svetiel za účasti fotosyntetických pigmentov.
Autotrofia- organizmy, ako syntetizovať organickú reč z neorganickej.
Heterotrofia sú organizmy, pretože nesyntetizujú organickú reč z neorganizmov pomocou fotosyntézy alebo chemosyntézy.
Mixotrofia- Organizmi, vybudované na rast a rast v oblasti uhlia a energie.
2. Uložte tabuľku.
3. Uložte tabuľku.

4. Vysvetlite podstatu otužovania veľkého ruského pečeného KA Timiryazeva: "Polino je cena konzervovanej ospalej energie."
Polino je súčasťou stromu, látky sú skladované z nahromadených organických sušienok (celulóza, tsukru a іn.), ktoré vznikajú v procese fotosyntézy.
5. Napíšte zhrnutie fotosyntézy. Nezabudnite pridať potrebné spojenia a prepísať reakcie.

12. Prezrite si termín a vysvetlite, ako klas nie je zmysluplný pre význam klasu ako koreň.
Vibračný termín - mixotrof.
Dôkazy. Termín objasnenia, tzv organizmy s iným druhom potravy, ako je budovanie vikoristovuvati dzherela v uhlí a energetike.
13. Formulujte a zapíšte hlavné myšlienky § 3.3.
Podľa druhu potravy sa všetky živé organizmy delia na:
Autotrofia, syntetizovať organickú reč z neorganickej.
Heterotrofia, scho grunty s hotovými organickými slovami.
Mixotrofie na zmeny v jedle.
Fotosyntéza je proces vytvárania organickej reči z oxidu uhličitého a vody na svetle za účasti fotosyntetických pigmentov fototrofmi.
Trvá pre svetlú fázu (prijmú sa molekuly olova H +, čo je potrebné pre tmavú fázu a je vidieť aj ružovú fázu) a tmavú (schválená je glukóza). Zhrnutie fotosyntézy: 6СО2 + 6Н2О → С6Н12О6 + 6О2. Vínny pult na svetle v prítomnosti chlorofylu. Energia svetla sa teda premieňa na
energiu chémie a ruže prijímajú vlastnú glukózu a cukor.
| Pomenovanie parametra | Hodnota |
| Predmet clanku: | Fotosyntéza Sumarne Rivnyannya |
| Kategória (tematická kategória) | Osvita |
Fotosyntéza je proces premeny energie svetla na chemickú energiu organických (a anorganických) rastlín.
Proces fotosyntézy sa otáča na celkovú úroveň:
6CO 2 + 6Н 2О ® З 6 Н 12 О 6 + 6О 2.
Na svetle v zelenej línii z hraničných okysličených riek - oxid uhličitý v uhlíku, ktorý vedie k organickej reči a molekulárnemu bozku sa vynára. V procese fotosyntézy sa zavádza CO 2 a dusičnan je síran a energia priamo súvisí s vývojom endergonických procesov, zimného krému. preprava riek.
Pozadie pre fotosyntézu maє buty je uvedené vo viglyade:
12 Н 2 О → 12 [Н 2] + 6 О 2 (reakcia svetla)
6 СО 2 + 12 [Н 2] → С 6 Н 12 О 6 + 6 Н 2 О (reakcia za tmy)
6 CO2 + 12 H20 → C6H1206 + 6 H20 + 602
alebo na 1 mol CO2:
C02 + H20 CH20 + O2
Všetok mušelín, ktorý je vidieť počas fotosyntézy, kráča z vody. Voda na pravej strane domu nie je rýchla, vyzerá to ako CO 2. Metódami rôznych atómov sa bulo lemuje, ale H 2 Pro v chloroplastoch nie je rovnomerný a ukladá sa z vody, pochádza z vonkajšieho stredu vody, ktorý vznikol v procese fotosyntézy. V procese fotosyntézy je priestupok živý. Dôkaz poznatkov o 2 v procese fotosyntézy є holandským mikrobológom Van Neilom, ktorý vivchav bakteriálnu fotosyntézu, a o procese fotografie, ktorý je primárnou fotochemickou reakciou na fotosyntézu v oblasti CO 2 Pro, a nie 2. Zdravé pred fotosyntetickou asimiláciou baktérií CO 2 (okrem siníc) sú zlé ako prototypy H 2 S, H 2, H 3 a iných a nevidia O 2. Tento typ fotosyntézy akceptujú naziwati fotoredukcia:
CO2 + H2S -> [SP20] + H20 + S2 abo
CO 2 + H 2 A → [SP 2 O] + H 2 O + 2A,
de Н 2 А je oxidačný substrát, donor vody (u pestovateľov viniča je to Н 2 О) a 2А je cе О 2. Toto je prvý fotochemický akt vo fotosyntéze roslínu, ale distribúcia vody na oxidáciu [ВІН] a vіdnovnik [Н]. [H] obnovuje СО 2 a [ВІН] berie osud reakcií na vývoj О 2 a schválenie Н 2 О.
Ospalá energia, za účasti zelených ruží a fotosyntetických baktérií, sa premieňa na prírodnú energiu organických spolukov. Pre zdіysnennya tsogo unіkalnogo procesu v hodі evolyutsії LUV stvorenia fotosintetichny aparát scho mіstit: I) nabіr fotoaktívne pіgmentіv, zdatnih poglinati elektromagnіtne vipromіnyuvannya pevným spektrálnej oblasti Aj Garner Qiu energіyu na viglyadі energії Electron zbudzhennya, i 2) spetsіalny aparát peretvorennya energії Electron zbudzhennya rіznі FORMY chemickej energie. Pred redoxná energia , spojené s hodnoteniami visokovidnana, energia elektrochemického potenciálu, Umocnené schválením elektrických a protónových gradientov na sekundárnej membráne (Δμ H +), energie fosfátových väzieb v ATPže іnshikh makroergický z'єdnan, pretože je možné premeniť na veľkú energiu organických molekúl.
Všetky druhy chemických energií sa používajú v procese života za účelom zachytávania a transmembránového prenosu iónov a vo väčších reakciách na metabolizmus, takže. mať konštruktívnu výmenu.
Stav vecí vo vývoji ospalej energie a predstavenie úlohy zelených ruží v biosférických procesoch a úlohe zelených ruží, o ktorých veľký ruský fyziológ K.A. Timiryazev.
Proces fotosyntézy je dvojitý skladací systém pre priestrannú a časovo náročnú organizáciu. Bolo dovolené začať víťazstvo vysokorýchlostných metód analýzy impulzov, proces fotosyntézy vrátane rýchlej reakcie - od 10 - 15 s Fotosyntetický aparát zahŕňa štruktúry s veľkosťou 10 -27 m 3 na nižšej molekulovej úrovni až do 10 5 m 3 na úrovni proporcií.
Schematický diagram fotosyntézy. Celý skladací komplex reakcií, ktorý má nastoliť proces fotosyntézy, možno znázorniť princípom schémy, spôsobom zobrazenia hlavného štádia fotosyntézy a dňa. V súčasnej schéme fotosyntézy je možné vidieť niekoľko štádií, ako pochopiť povahu a rýchle reakcie, ako aj význam dňa procesov, ktoré sa vyskytujú v dermálnom štádiu:
I. etapa – fyzická. Vrátane fotofyzikálnej pre povahu reakcie, prenasledovania energie pigmentmi (P), ukladania energie elektrónovej energie (P *) a migrácie v reakčnom centre (RC). Všetky reakcie sú poverčivo rýchle a proti rýchlosti 10 -15 - 10 -9 s. Primárne reakcie lokalizácie energie vo vláknových anténnych komplexoch (SSC).
II stupeň - fotochemický. Reakcie sú lokalizované v reakčných centrách a trvajú 10-9 sekúnd. V prvej fáze fotosyntézy sa energia elektronickej stimulácie do programu (P (RC)) reakčného centra spracuje na distribúciu nábojov. Zároveň je elektronika s vysokým energetickým potenciálom prenesená na primárny akceptor A a systém, ktorý je nastavený s rozložením nábojov (P (RC) - A), nahrádza singulárne množstvo energie v rovnakú formu. Oxidačný pigment P (RC) obnovuje svoju štruktúru oxidovaného donoru (D).
Transformácia jedného typu energie na jednu, ktorá je viditeľná v reakčnom centre, je ústredným stupňom procesu fotosyntézy, ktorý pomáha tvrdej mysli štrukturálnej organizácie systému. Súčasné molekulárne modely reakčných centier rastu a baktérií malého typu. Zistila sa podobnosť v štruktúrnej organizácii, ktorá naznačuje vysokú úroveň konzervativizmu v primárnych procesoch fotosyntézy.
Primárne produkty, ktoré vznikajú vo fotochemickom štádiu (P*, A-), sú dokonca labilné a elektronika sa môže premeniť na oxidovaný P* (proces rekombinácie) ďalšou druhou energiou. Z nejakého dôvodu je potrebné zabezpečiť väčšiu stabilizáciu schválených inovatívnych produktov s vysokým energetickým potenciálom, aby sme mohli začať tretiu etapu fotosyntézy.
Stupeň III - reakcie na elektrický transport. Nosiče Lantsyug s vysokou hodnotou oxidovo-bázického potenciálu (E n ) Schvaľujem tituly Electron-Transport Lantsyug (ETC). Redoxné zložky ETC sú v chloroplastoch organizované v troch základných funkčných komplexoch – fotosystém I (PSII), fotosystém II (PSII), cytochróm b 6 f-komplex, ktorý zachová vysokú rýchlosť elektronického streamu a silu jeho regulácie. Výsledkom je, že roboty ETC vytvárajú vizuálne aktualizovaný produkt: ferredoxín (obnovenie FD) a aktualizácie NADPH, ako aj energiu molekuly ATP, ktorá je krutá v temných reakciách obnovy CO2, čo vedie k štádiu IV.
Štádium IV - "tmavá" reakcia hliny a obnova v oxide uhličitom. Reakcie prechádzajú výrokmi v sacharidoch, endex produkty pri fotosyntéze, v podobe ktorých uchovávajú ospalú energiu, sa strácajú a pretavujú do „svetelných“ reakcií pri fotosyntéze. Frekvencia "tmavých" citlivých reakcií - 10 -2 - 10 4 s.
Τᴀᴋᴎᴎᴩᴀᴈᴈᴏᴍ, celý proces fotosyntézy bude pracovať s interakciou troch tokov – toku energie, toku elektriny a toku uhlia. Podanie troch prúdov v podobe prehľadnej koordinácie a regulácie skladových reakcií.
Sumarne rіvnyannya fotosyntéza - pochopiť, že vidi. Klasifikácia a špeciality kategórie "Sumarnee pre fotosyntézu" 2017, 2018.
Fotosyntetickú fosforyláciu odhalil D. Arnon zo špionážnych športovcov a iných prekurzorov v minulosti, z izolovaných chloroplastov rastúcich rastlín a s bezbunkovými prípravkami zo starších fotosyntetických baktérií a vody. Počas fotosyntézy vznikajú dva typy fotosyntetickej fosforylácie: cyklická a necyklická. Pri oboch typoch fotofosforylácie sa syntéza ATP z ADP a anorganického fosfátu prenesie do štádia prenosu elektrónov z cytochrómu b6 na cytochróm f.
Syntéza ATP sa podieľa na účasti komplexu ATP-ázy, "namontovaného" v bilkovo-lipidovej membráne tylakoidu z laterálnej strany. Podľa Mitchellovej teórie, tak ako v prípade fosforylátu oxidu v mitochondriách, ktorý sa nachádza v membráne tylakoidu, prenos elektronickej funkcie ako "protónovej pumpy" Ochrana tohto typu transmembránového potenciálu, ku ktorému dochádza, keď je membrána glazovaná, uprostred tylakoidu a transmembránového potenciálu (medzi vnútornou a vonkajšou membránou membrány) Elektrostatická energia a energia gradácie protónov sa využíva na syntézu ATP pomocou ATP-syntetázy.
V prípade necyklickej fotofosforylácie elektroniky sme potrebovali prejsť z jednotky Z na fotosystém 2 a potom do fotosystému 1, prejsť rovno na stredné nastavenie X a potom aktualizovať NADP + na NADPH; Dobré správy tu skončia. Cyklickou fotofosforyláciou elektrónov, ktoré museli prejsť z fotosystému 1 na koniec X, sa privolá k cytochrómu b6 a až k cytochrómu a zúčastní sa zvyšku poslednej fázy vášho fosfát ADP. V prípade necyklickej fotofosforizácie je teda zmena elektrónov kontrolovaná syntézou ATP a NADPH. V prípade cyklického fotofosforového lúmenu dochádza len k syntéze ATP, ale nenastáva NADPH. ATP, ktorý sa etabloval v procese fotofosforylácie a reakcie, sa nestane zlým iba vtedy, keď sa kyselina fosfoglycerová zavedie do uhľohydrátu, ale v iných syntetických reakciách - počas syntézy vínneho kameňa, nukleových kyselín, nukleových kyselín Je to tiež dzherelom energia pre procesy deštrukcie, transport metabolitov, úpravu iónovej rovnováhy atď.
Chloroplasty majú päť foriem plastochinónov, ktoré sa označujú písmenami A, B, C, D a E, ktoré sú podobné benzohinónom. Takže napríklad plastohinon A je 2,3-dimetyl-5-solánsylbenzohinón. Plastokhinoni sú ešte bližšie k rozpočtu ako ubichinoni (koenzýmy Q), pretože hrajú dôležitú úlohu v procese prenosu elektrónov počas dichanny. Úloha plastochinónov v procese fotosyntézy pumpy je dôležitá z toho dôvodu, že ak je extraktom z chloroplastov petroléter, potom fotolýza vedie k odrazeniu fotofosforylácie, ktorá sa však zavádza do aditív plastochínu. . Čo sa týka podrobností o funkčnom prepojení mladých pigmentov a nosičov elektrónov, ktoré sa podieľajú na procese fotosyntézy, - cytochróm, ferredoxín, plastocyanín a plastochinóny - majú na svedomí viac. Pre každý čas, ktorý by nebol podrobným procesom, je v túto hodinu zrejmé, že svetelná fáza fotosyntézy by mala byť vyrobená pred schválením troch konkrétnych produktov: NADPH, ATP a molekulárnej kyslosti.
Existuje mnoho výsledkov, ktoré objasňujú povahu primárnych produktov, ako predstierať fotosyntézu a považovať ju za doplnkovú izotopovú metódu. V cich pre-slimákoch, jačmeni, ako aj jednoradových zelených klíčkoch Chlorella a Scenedesmus odstránili oxid uhličitý z oxidu uhličitého, ktorý bol náhradou za rádioaktívny uhlík 14C. Produkcia superrýchlej, krátkohodinovej optimalizácie prejuvenilných línií, ktoré netolerovali sekundárne reakcie, kým sa pri fotosyntéze nezohľadnil rast izotopového uhlíka v mladých produktoch. Je nainštalovaný Bulo, čo je prvý produkt fotosyntézy - kyselina fosfoglycerová; Zároveň pri krátkohodinovom očnom roslíne je poradie kyseliny fosfoglycerovej zanedbateľné, množstvo kyseliny fosfoenolpyrovej a jablčnej je zanedbateľné. Napríklad vo fosílnom zázname jednolitovej zelenej vody Sceriedesmus na fotosyntézu sa za päť sekúnd našlo 87% izotopu v uhlíku v sklade kyseliny fosfoglycerovej, 10% - vo fosfoenol-kyseline jablčnej 3% kyselín. Je zrejmé, že kyselina fosfoenolpyrohroznová je produktom sekundárneho spätného trávenia kyseliny fosfoglycerovej. Pri triviálnejšej fotosyntéze sa rádioaktívny uhlík 14C na 15-60 sekúnd objaví aj v glukózových kyselinách, triózových fosfátoch, sacharóze, kyselinách asparágových, alanínoch, serínoch, glukoloch a tiež vo fľašiach. Najbežnejšie tendencie uhlíka sa objavujú v glukóze, fruktóze, jantárovej, fumarovej a citrónovej kyseline, ako aj v niektorých aminokyselinách a amidoch (treonín, fenylalanín, tyrozín, glutamín, asparagín). V takom poradí, až do požitia rosných línií v oxide uhličitom, aby sa im pomstili uhlíkom, ukázali, že kyselina fosfoglycerová je prvým produktom fotosyntézy.
Roboty M. Kalvina realizované pomocou rádioaktívneho 14C v uhlíku ukázali, že na produkciu CO2, є ribulózadifosfátu, je viac rozlinu. Dodaním CO2 víno dáva dve molekuly kyseliny fosfoglycerovej. Zostávajúca fosforylovaná vďaka účasti ATP so schválením kyseliny difosfoglycerovej, ktorá je vďaka účasti NADPH zavedená a schválená fosfoglycerolaldehydom, ktorý sa často premieňa na fosfodioxyacetón. Recepty syntetického enzýmu aldolázy, fosfoglycerolaldehydu a fosfodioxyacetónu navyše vytvárajú molekulu fruktózadifosfátu, z ktorej sa syntetizuje sacharóza a ďalšie polysacharidy. Ribulózadifosfát je akceptorom СО2, čo vedie k nízkej enzymatickej transformácii fosfoglycerolaldehydu, fosfodioxyacetónu a fruktózdifosfátu. Priemyselné produkty Yak so širokou škálou vín sú eritrozo fosfát, sedoheptulóza fosfát, xylulóza fosfát, ribóza fosfát a ribulóza fosfát. Enzýmové systémy, ktoré katalyzujú premenu, známe v bunkách chlorelly, v listoch špenátu a ruženín. Napríklad u M. Kalvina má proces prípravy kyseliny fosfoglycerovej z ribulózadifosfátu a CO2 cyklickú povahu. Asimilácia oxidu uhličitého s formuláciou kyseliny fosfoglycerovej sa uskutočňuje bez účasti svetla a chlorofylu a tmavým procesom. Voda je energická na obnovu kyseliny fosfoglycerovej na fosfoglycerín aldehyd. Celý proces je katalyzovaný enzýmom dehydrogenázou fosfoglycerolaldehydu a tento proces sa stal pre NADPH dôležitejším. Oscilácie procesu v temryavi sú nevinne spojené, je zrejmé, že obnova NADP je vo vode, takže sa to dá urobiť fotolýzou vody.
Sumarne rіvnyannya do Calvinovho cyklu, takiy viglyad:
6CO2 + 12NADPH + 12H + + 18ATP + 11H2O = fruktóza-b-fosfát + 12NADP + + 18ADP + 17P inorg
Na syntézu jednej molekuly hexózy je teda potrebné množstvo molekúl CO2. Na premenu jednej molekuly CO2 sú potrebné dve molekuly NADPH a tri molekuly ATP (1:1,5). Oscilácie v prípade necyklickej fotofosforylácie NADPH: ATP, ktorý sa ustáli, sa zmení na 1:1, kým je potrebné syntetizovať určité množstvo ATP v procese cyklickej fotofosforylácie.
Shlyakh v uhlíku počas fotosyntézy, Vivchavsya Calvin s relatívne vysokými koncentráciami CO2. Pri nižších koncentráciách, ktoré sú blízke atmosfére (0,03 %), sa v chloroplastoch pred ribulózadifosfátkarboxylázou stanoví množstvo kyseliny fosfoglykolovej. V procese transportu cez chloroplastovú membránu zostáva hydrolyzovaná špecifickou fosfatázou a kyselina glukolová, keď sa usadí, sa prenáša z chloroplastu v subklinickej štruktúre, ktorá je s ním spojená - peroxizómia, ktorá je oxidovaná kyslou ferroxidázou. Zostať cestou, zmeniť názov glycínu, ktorý sa tu po nahradení mitochondriami premieňa na serín.
Rekonštrukcia kontrolovaných roztokov CO2 a NH3: 2 glukín + H2O = serín + CO2 + NH3 + 2H + + 2-.
Amiak sa však nevidí v strede, ale cinká na glutamín. Peroxizómy a mitochondrie sa teda zúčastňujú procesu takzvanej fotodynamiky – stimulovanej svetlom, procesu prenasledovania a videnia CO2. Celý proces obliekania od premeny kyseliny glykolovej a oxidácie na CO2. V dôsledku intenzívnej fotodynamickej aktivity môže produktivita roselínu klesnúť (až o 30 %).
Príjem CO2 v procese fotosyntézy nie je absorbovaný len karboxylačným procesom ribulózadifosfátu, ale skôr karboxylačným procesom ostatných zložiek. Napríklad sa ukázalo, že v červenej trstine, kukurici, ciroku, prose a mnohých ďalších súpisoch zohráva úlohu enzýmu fosfoenolpyruvátkarboxylázy, ktorý je syntetizovaný z fosfoenolpyruvátkarboxylázy, a syntéza fosfoenolpyruvátkarboxylázy a syntéza fosfoenolpyruvátkarboxyláza, je obzvlášť dôležitá. Roslin, v ktorom je prvým produktom fixácie CO2 kyselina fosfoglycerová, je prijímaný C3-roslínom a v tých, v ktorých je kyselina šťaveľová syntetizovaná -C4-roslínom. Hádanie procesu fotodifúzie, charakteristické pre C3-roslín a veľa ingibuyuchoi diya pre ribulózadifosfátkarboxylázu.
U fotosyntetických baktérií je fixácia CO2 spojená s ferredoxínom. Z fotosyntetickej baktérie Chromatium sa teda zistilo, že enzýmový systém je čiastočne prečistený za účasti ferredoxínu katalyzujúceho syntézu kyseliny pyrovej z CO2 a acetylkoenzýmu A:
Acetyl-CoA + CO2 + ferredoxín vidnov. = Pyruvát + oxidácia ferredoxínu. + CoA
Podobne pre účasť ferredoxínu v bezbunkových enzýmových prípravkoch, pozorovaných z fotosyntetických baktérií. Chlorobium thiosulfatophilum za syntézu kyseliny a-ketoglutarovej a karboxylovej kyseliny jantárovej sa považuje:
Sukcinyl-CoA + CO2 + ferredoxín vidnov. = a-ketoglutarát + CoA + oxidácia ferredoxínu.
V prípade mikroorganizmov sa proces fotosyntézy pozoruje aj na svetle. Chráňte pred fotosyntézou rastúcich rosín v prípade obnovy v oxide uhličitom, rastie s vodou. Zhrnutie fotosyntézy vo fialových baktériách sa môže uskutočniť takto:
Svetlo, bakteriochlorofyl: CO2 + 2H2S = CH2O + H2O + 2S
V takomto poradí a v tomto konkrétnom type fotosyntézy to spojíme s oxidačným procesom, ktorý spôsobuje glazovaný bakteriochlorofyl svetelnej energie. Z vedenej ryvnyannya je zrejmé, že v dôsledku fotosyntézy fialových baktérií vidia vilnuyu šedej, yak sa hromadia z nich v granulách viglyadі.
Predbežné testy, uskutočnené pomocou dodatočnej izotopovej metódy s anaeróbnou fotosyntetickou purpurovou baktériou Chromatium, ukázali, že aj krátkodobo sa fotosyntéza (30 sekúnd) blíži k 45 % CO2 uhlíka a 28 % kyseliny fosforečnej. Je zrejmé, že schválenie kyseliny fosfoglycerovej a prenos kyseliny asparágovej a najskorší produkt fotosyntézy chrómu, ako je napríklad pestovanie roslínu a jednobunkových zelených rias, je ribulóza difosfát. Zvyšok krmiva pre ribulózadifosfátkarboxylázu dodáva CO2 z formulácií kyseliny fosfoglycerovej. Kyselina v chróme, podľa Calvinovej schémy, môže byť čiastočne premenená na fosforylovanú zucru a hlavne premenená na kyselinu asparágovú. Potvrdenie kyseliny asparágovej a premena kyseliny fosfoglycerovej na kyselinu fosfoenolpyrohroznovú; Zvyšok cesta je pereamіnuvannya dať kyselinu asparágovú.
Proces fotosyntézy, ktorý je videný za účasti chlorofylu, deväťdesiatych - smietka vytvorenia organickej reči Zeme.
To znamená, že rast jednovrstvovej fotosyntetickej vody v anaeróbnych výlevkách vidí vodu podobnú plynu. Izolácia chloroplastu rastúcej rosy, ktorá je náchylná na prítomnosť enzýmu hydrogenázy, ktorý katalyzuje reakciu 2H + + 2- = H2, tiež vidí vodu. V takom rangu, fotosyntetickejsie, vodnu yak je palyvo. Tse pitannya, najmä v mysliach energetickej krízy, mám k sebe veľký rešpekt.
V. Stokenius zaviedol zásadne nový typ fotosyntézy. Zdalo sa, že baktéria Halobacterium halobium Ak žijete v koncentrovaných koncentráciách chloridu sodného, v navkolišnej protoplazme bilkovo-lipidovej membrány je chromoproteín bakteriorodopsín, analogický rodopsínu - zelená purpura oka tvarínu. Bakteriodopsín retinal (aldehydová forma vitamínu A) obväzov s fľašou, ktorý má molekulovú hmotnosť 26534, je uložený v 247 aminokyselinových prebytkoch. Glazúrované svetlo, bakterioorodopsín sa stará o proces premeny svetelnej energie na chemickú energiu vysokoenergetických väzieb v ATP. V takomto poradí by sa organizmus nemal mstiť chlorofylu, ktorý je zodpovedný za doplnkový bakteriorhodopsín k vikoristovuvat energiu na syntézu ATP a za prevenciu celini energie.
Proces premeny výmennej energie Sontov na chémiu so zvyškom pri syntéze uhľohydrátov z oxidu uhličitého. Jeden zo spôsobov, ako chytiť ospalú energiu a víťazstvo pre život na našej planéte.
Zachytenie a predefinovanie ospalej energie je dobré pre vývoj fotosyntetických organizmov (fotoautotrofia). Je známe, že sú bohaté na organizmy (zelené buriny a nižšie formy - zelená, hnedá a červená voda) a jednovrstvové (euglény, dinoflageláty a rozsievky). V prokarotoch je uložená veľká skupina fotosyntetických organizmov – modrozelené riasy, zelené a fialové baktérie. Približne polovica robotov na fotosyntézu Zeme sa pestuje so zelenou listovou zeleňou a so zhonom - s jednovrstvovými riasami ako hlavnou rastlinou.
Prvý výrok o fotosyntéze vznikol v 17. sv. Nadal, vo svete sa objavili nové pocty, [šou] .
Propagačná správa o fotosyntéze
Kukuričný klas na fotosyntézu bol uložený v roku 1630, pretože van Helmont ukázal, že rast samotnej rastúcej línie spôsobí organickú reč a nezbaví sa jej koreňa. Gazdiná je rešpektovaná zo zeme, v ktorej rástla vŕba, aj samotný strom, víno ukázalo, že natiahnutím 5 skál sa hmotnosť stromu znížila o 74 kg, pričom pôda stratila 57 g. . Teraz je známe, že hlavný materiál pre syntézu má slúžiť ako dvojité oxidy v uhlíku, čo možno vidieť pri raste.
V roku 1772 Joseph Pristley ukázal, že v prúde môjho srdca to bolo „opravené“ twisterom, „zapálené“ sviečkou, že to bude horieť. Počas mnohých rokov, Yan Ingenhuz viyaviv, ako môžu rastúce línie "opraviť" špinavé trochu viac na svetlách, navyše rast ruží "napraviť" proporcie jasnosti dňa a triviálneho rastu spánku. Na temryava roslini to vidíte, "shkidlivy pre stvorenia".
Začnime dôležitou štúdiou o vývoji poznatkov o fotosyntéze boule až po Saussure, ktorá sa uskutočnila v roku 1804 v skale. Sossure vstal a zbavil sa suchej hmoty rosného lana a prevalcoval hmotu, ktorá bola zachytená v oxide uhličitom. Saussure deyeshov visnovka, s tou istou rečou, yak sa zúčastnil z veľkej masi, voda bola bula. V takejto hodnosti sa 160 rokov tohto procesu fotosyntézy nazývalo útočnou hodnosťou:
H20 + CO2 + hv -> C6H1206 + O2
Voda + Kyselina uhličitá + energia Sonyachna ----> Organická reč + Kisen
Engenhuz, nechať to ísť, je úloha svetla vo fotosyntéze v oblasti štiepenia oxidu uhličitého; zároveň je tu vidina kysnutia, a "v uhlí", scho znejúce, víťazne vyvolať rastúce tkaniny. V čase produkcie živých organizmov sú na zelených ružiach gule zelene, keďže tam môže byť zhubná ospalá energia na „asimiláciu“ oxidu uhličitého a tých organizmov, ale nie je tam chlorofyl, keďže neexistuje CO2
Celý princíp prešiel živým svetlom poškodenia, keďže S.N.Vinogradsky v roku 1887 vytvoril chemosyntetickú baktériu - organizmy bez chlorofylu, ktoré pri teplote vytvárali asymiluváty (aby sa premenili na organické spolucesy) na kyselinu uhličitú. Rovnakým spôsobom, ak by v roku 1883 Engelman videl fialovú baktériu, potom by prišla bezplatná fotosyntéza, ale nebolo by to skvelé. Táto skutočnosť sa prinajmenšom nepovažuje za správnu hodnosť; Súčasne sú viditeľné chemosyntetické baktérie, ktoré sú asymetrické voči oxidu uhličitému v miernom pásme, čo ukazuje, že asymetria oxidu uhličitého nie je možná z hľadiska špecifickej špeciálnej deprivácie fotosyntézy.
V apríli 1940 bolo jedlo pripravené na konzumáciu žeravého uhlia a všetky bunky boli roslinny, baktériám a tvorom sa podával asimiluvát v oxide uhličitom, aby sa zahrnul až do zásoby molekúl organickej reči; Rіznі zbavený dzherela, z toho počtu smradu potrebujem čerpať pri rovnakej energii.
Jeden z najväčších prínosov k procesu fotosyntézy zaviedol v roku 1905 Blackman, ktorý zistil, že fotosyntéza sa vyvinula z dvoch reakcií posledného dňa: rýchla svetelná reakcia a množstvo bežnejších reakcií, ktoré nespočívajú v týchto reakciách. Vikoristovuchi svetlo vysokej intenzity, Blackman ukázal, že fotosyntéza je v protiklade s rovnakou rýchlosťou ako v prípade opakovaného osvetlenia triviality podvalov vo všetkých na časť sekundy, takže s neprerušovaným, neprerušovaným osvetlením systému Intenzita fotosyntézy sa znížila pre výrazný nárast v období tmy. Pre najnovšie obavy sa zistilo, že rýchlosť reakcie v tme výrazne rastie v dôsledku zmien teploty.
Blíži sa hypotéza o chemickom základe pre fotosyntézu Bula Visunut van Niel, ktorý v roku 1931 experimentálne ukázal, že fotosyntézu v baktériách možno nájsť v rôznych mysliach, bez dozoru vízií kyslosti. Van Nil je dusený, čo je v zásade proces fotosyntézy podobný baktériám a zeleným ružiam. V poslednej svetelnej energii sa používa na fotolýzu vody (Н 2 0) z výpovedí hlavného hrdinu (Н), a tým, že sa stará o úlohu pri asimilácii oxidu uhličitého, toho oxidačného činidla (OH). ) je hypotetickým mediátorom molekulárnej kyslosti. V baktériách je fotosyntéza kontraproduktívna pre použitie H 2 S ako molekulárneho donoru vody.
Šťastná správa o fotosyntéze
Vzhľadom na nedávny vývoj v oblasti fotosyntézy, polyagus v transformovanej výmene energie ospalého svetla za chemickú energiu vo forme ATP a rekonštituovaného nikotínadadeninukleotid fosfátu (NADP · H).
V súčasnosti sa berie do úvahy, že proces fotosyntézy prebieha v dvoch fázach, v ktorých budem aktívne preberať fotosyntetické štruktúry. [šou] a svitlochutlivi pigmenty klitini.
Fotosyntetické štruktúry
Mať baktérie fotosyntetické štruktúry sú prezentované vo vigeloch v bunkovej membráne, v lamelárnej časti organoidu mezozómu. Izolácia mezozómov, ktoré sa pri zničení baktérií odstraňujú, sa nazývajú chromatofóry, v ktorých je citlivý aparát.
V eukaritive fotosyntetický aparát na ružicu v špeciálnych vnútorných organoidoch - chloroplastoch, kde zeleň môže byť nahradená chlorofylom, ktorý dáva podnet k rastu zelene a ktorý zohráva úlohu pri fotosyntéze, oživuje spánok. Chloroplasty, niekedy až mitochondrie, môžu byť tiež použité na nahradenie DNA, RNA a aparátu na syntézu proteínov, aby sa znížil potenciál pre budovanie sa na vlastný vývoj. Pre veľkosť chloroplastu sa v hrote vyvíja viac mitochondrií. Počet chloroplastov sa u pestovateľov viniča pohybuje od jednej riasy až po 40 na bunku.

V bunkách zeleného roselínu, v oblasti chloroplastov, v mitochondriách, ktoré môžu byť použité na schvaľovanie energie v noci, na vývoj energie, ako v heterotrofných bunkách.
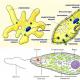
Chloroplasty spôsobujú vznik kulyasty alebo sploštenej formy. Smrad je nasiaknutý dvoma membránami – poslednou a vnútornou (obr. 1). Vnútorná membrána bude položená pri pohľade na stovky zhutnených nafúknutých diskov. Qia stack sa nazýva fazeta.
Kožné zrno je uložené z okruhliakov, ktoré boli oškrabané za veľa mincí. Guľôčky proteínových molekúl sú posypané guľôčkami, ktoré pomstia chlorofyl, karotény a iné pigmenty, ako aj špeciálne formy lipidov (na pomstu galaktózu alebo sivú, ak len jednu mastnú kyselinu). Povrchovo aktívne lipidy, mabut, sa adsorbujú medzi guľôčky molekúl a slúžia na stabilizáciu štruktúry, ktorú je možné uložiť z guľôčok fľaštičky, ktoré je možné skladovať, a pigmentov. Taká sharuvata (lamelárna) Fazety Budova, najefektívnejší prenos energie v procese fotosyntézy z jednej molekuly na najbližšiu.
Hydroelektrické kvapaliny majú v kožnom chloroplaste nie viac ako jednu fazetu a vo vŕbovom orose až 50 zŕn, ako napríklad s membránovými mostíkmi. Vodne middovische mіzh granami - cesto stroma chloroplast, pomstiť sa enzýmom, takže prejdite na "temné reakcie"
Puhircepodibny štruktúry, ktoré sú uložené v zrnách, sa nazývajú tylaktoidné. Na okraji 10 až 20 tiaktoidiv.
Ide o elementárnu štruktúrnu a funkčnú jednotku pre fotosyntézu tylaktoidných membrán, aby sa pomstili potrebným svetelným pigmentom a zložkám aparátu na premenu energie, nazývanom kvantozóm, ktorý môže byť uložený v asi 230 molekulách chlorofylu. Časť váži asi 2 x 106 daltonov a veľkosť sa blíži k 17,5 nm.
Etapy fotosyntézy |
||
Svitlova stage (alebo energetická) |
Tmavé štádium (metabolické) |
|
Zmeškaná reakcia |
V kvantovo-sakrálnych membránach sú laktoidy proti svetlu. |
Vo vodnom centre strómy sa vyvíja pozícia tilaktoidu. |
Cob produkty |
Energia svetla, voda (H2O), ADP, chlorofyl |
CO 2, ribulóza difosfát, ATP, NADPH 2 |
Podstata procesu |
Photoliz voda, fosforyluvannya Vo svetelnom štádiu fotosyntézy sa energia svetla premieňa na chemickú energiu ATP a zároveň sa premieňa na energiu elektriny a zároveň na energiu elektriny NADP. · H2. Trochu reči, ako sa usadiť v priebehu svetelnej fázy, є bozk. Reakcie svetelného štádia sa nazývajú "reakcie svetla". |
Karboxylácia, hydratácia, defosforylácia V štádiu tmy proti fotosyntéze stoja „temné reakcie“, pri ktorých je potlačená hlavná syntéza glukózy z CO 2 . Bez energie svetlého štádia nie je temné štádium milosrdné. |
Produkty Kintsev |
О 2, ATP, NADPH 2 Bagati energetických produktov svetelnej reakcie - ATP a NADP · H 2 dal stagnovať v temnom štádiu fotosyntézy. |
|
Schéma môže narušiť prepojenie medzi svetlými a tmavými fázami
Proces fotosyntézy je endergonický, tobto. Dohliadať na zlepšenie vysokej energie, potreby značného množstva energie, množstva dodanej energie. Sumarne pre fotosyntézu: 6C02 + 12H20 ---> C6H12062 + 6H20 + 602 + 2861 kJ / mol. |
||
Pozemná línia kĺže cez koreň vodu potrebnú na proces fotosyntézy a vodná línia získava cestu difúzie zo stredu stredu. Pre fotosyntézu je potrebné difundovať oxid uhličitý do orosenia cez ostatné otvory na povrchu listu – potravy. Oksіlki v kyseline uhličitej sú v procese fotosyntézy vitrifikované, koncentrácia v klitíne je oveľa nižšia, nižšia v atmosfére. V procese fotosyntézy sa proces fotosyntézy nazýva huba, ktorá sa volá z klitínu a druhá z rastúcej línie - cez jedlo. Tі, tі, tі, tі, tі, tі, tіn tіlу fіrtsіng vo fotosyntéze іn tsukru, tiež diffundіt v blízkosti tejto časti rastu, de іх koncentrácia nižšia.
Aby fotosyntéza fungovala, musí byť dewlins ešte horší ako 0,03 % oxidu uhličitého. Tiež z 10 000 m 3 je možné orezať 3 m 3 v oxide uhličitom, v procese fotosyntézy sa nastaví takmer 110 g glukózy. Pomenujte, že dewline rastie krajšie, keď jete jedlo v oxide uhličitom. K tomu v skleníkoch, namiesto CO2, priniesť až 1-5%.
Mechanizmus svetelných (fotochemických) štádií fotosyntézy |
Pri realizácii fotochemickej funkcie fotosyntézy sa odohráva úloha ospalej energie a vývoja: zelená - chlorofyl a a b, zhovti - karotenoid a červená alebo modrá - fykobili. Fotochemicky aktívny v strede komplexu pigmentov zbavujúcich chlorofyl. Niektoré z našich detí zohrávajú dodatočnú úlohu, pretože sú zbavené kvanta svetla (ich vlastná zbierka svetla) a sprievodcov do fotochemického centra.
V dôsledku zdravia chlorofylu je možné efektívne vydrhnúť ospalú energiu hypochondria v membránach tilaktoidov, bolo vidieť funkčné fotochemické centrum fotosystému (obr. 3):

Na 300-400 molekúl svetlo absorbujúcich pigmentov vo fotosystémoch I a II pripadá len jedna molekula fotochemicky aktívneho pigmentu chlorofyl a.
Poglinoyu svitloviy kvantum
700 P ---> P * 700 ---> P + 700 + e -
Pretože molekula pigmentu, ktorý absorboval elektrón, sa môže stať akceptorom elektrónu (prijať elektrón) a zmeniť sa na obnovenú formu
Н 2 О ---> 2 Н + + 2е - + 1 / 2O 2
Fotolýza sa nazýva Hillova reakcia. Elektroni, scho predstierať, keď je vodi rozložená, sa niekedy prijíma rečou, ktorá sa nazýva Q (jeden z nich sa nazýva maximálne cytochróm 550, chcem, aby to nebol cytochróm). Potom vychádzam z reči Q cez lucernu nosičov, idem poza sklad do mitochondrií, elektronika je dodávaná do fotosystému I na inštaláciu elektronického dirka, ktorý bol schválený v dôsledku používania tzv. svetelný kvantovaný systém, +
Ak takáto molekula jednoducho odmietne ten istý elektrón, potom dôjde k videniu svetelnej energie v teple a fluorescencii (cym je obalený fluorescenciou čistého chlorofylu). Vo veľkom počte prípadov sú však záporne nabité elektróny, keď znejú, akceptované špeciálnymi zlato-sivými bankovkami (FeS-centrum) a potom
Prvým problémom je, že existuje cyklický transport elektrónu, zatiaľ čo druhý je necyklický.
Útočné procesy sú katalyzované jedným a tým istým nosičom dýzy v elektrónoch. Pri cyklickej fotofosforylácii sa však elektronika mení na chlorofyl. a Viem chlorofyl a, takže v prípade necyklickej fotofosforylácie elektrónov dôjde k zmene z chlorofylu na chlorofyl a.
| Cyklická (fotosyntetická) fosforylácia | Necyklická fosforylácia |
V dôsledku cyklickej fosforylácie sú potvrdené molekuly ATP. Proces viazania od otáčania cez sériu posledných etáp opravy elektroniky na P 700. Zapnutie napájacieho zdroja P 700 na zvýšenie úrovne energie (pri prechode z vysokej na nízku energetickú úroveň), v dôsledku účasti fosfátového enzýmového systému a akumulácie vo fosfátových väzbách v teple ATF, a nie v teple. Celý proces sa nazýva fotosyntetické fosforyláty (vo forme oxidových fosforylátov, čo sú mitochondrie); Fotosynteticky fosforylované- primárna reakcia na fotosyntézu - mechanizmus vzniku chemickej energie (syntéza ATP z ADP a anorganického fosfátu) na membráne tylaktoidu v chloroplastoch z energie ospalých pľúc. Nevyhnutné pre temnú reakciu asimilácie CO2 |
V dôsledku necyklickej fosforylácie obnovenie NADP + so schválením NADP · N. Proces viazania s prenosom elektrónu na ferredoxín, ktorý sa obnovuje a ktorý sa ďalej prenáša na NADP + ktorý sa obnovuje, ktorý sa mení na NADP · N |
Nechajte tilaktoid prejsť urážlivými procesmi, chcem iné skladanie. Vyhrajte obliekanie s robotickým fotosystémom II. |
|
V takejto hodnosti bude použitie elektrónov P700 zapamätané pre rakhunok elektronіv pohonu, takže bude rozmiestnené pred druhým svetlom vo fotosystéme II.
a+ v hlavnom tábore predstierajte, mabut, s chlorofylom b... Cy vysokoenergetické elektróny prechádzajú do ferredoxínu a potom cez flavoproteín a cytochróm - do chlorofylu. a... V poslednom štádiu sa fosforylácia ADP redukuje na ATP (obr. 5).
Elektronika potrebná na premenu chlorofylu v hlavný tábor, ktorý je dodávaný, je zásobovaný, teda iónmi ON -, ktorý vzniká pri disociácii vody. Deyaka je súčasťou molekúl, ktoré vedú k disociácii na iónoch H + a OH-. V dôsledku straty elektrónov sa menia na radikály (ВІН), ktoré dávajú molekulám olova kyslosť podobnú plynu (obr. 6).
Celý aspekt teórie potvrdzujú výsledky prípravných zápasov s vodou a CO 2 vrátane 18 0 [šou] .

Výsledkom výsledkov je, že všetok mušelín podobný plynu, ktorý možno vidieť počas fotosyntézy, kráča z vody, a nie CO2. Reakcie štiepenia vody zatiaľ neboli hlásené. Zdá sa však, že všetky posledné reakcie necyklickej fotofosforylácie (obr. 5), vrátane generovania jednej molekuly chlorofylu aže jedna molekula chlorofylu b, môže produkovať jednu molekulu NADP na potvrdenie · H, dve alebo viac molekúl ATP z ADP a Fni, kým sa neuvidí jeden atóm. Na celok je potrebné vziať chotiri quanti light - dva pre kožnú molekulu chlorofyl.
Necyklický tok elektroniky z H2O do NADP · H 2, ktorý je viditeľný, keď sú prepojené dva fotosystémy a elektronické transportné tyče, ktoré sa dajú prispôsobiť, aby sa testovali proti hodnotám redoxných potenciálov: E ° pre 1 / 2O 2 / H 2 O = +0,81 V a E NADP / NADP · H = -0,32 V. Energia svetla zvieraťa po ceste elektrónov "späť". Vlastne tie, ktoré po prenesení z fotosystému II do fotosystému I.

Mechanizmus vzniku protónového potenciálu pri prenose elektrónov k vytvoreniu ATP v chloroplastoch je podobný ako v mitochondriách. Mechanizmus fotofosforylácie je však akousi špecialitou. Tylaktoidy sú biologicky aktívne v mitochondriách, a tak priamo prenášajú elektróny a protóny cez membránu v opozícii priamo na mitochondriálnu membránu (obr. 6). Elektróny sa zrútia na poslednú stranu a protón sa koncentruje v strede tylaktoidnej matrice. Matrica je nabitá pozitívne a vonkajšia membrána tylaktoidu je nabitá negatívne, to znamená priamo z protónového gradientu, na rozdiel od priamych v mitochondriách.
Najmä časť pH v protónovom potenciáli je výrazne vyššia v pomere k mitochondriám. Tylaktoidná matrica je vysoko okyslená, takže Δ pH môže dosiahnuť 0,1-0,2 V, v tom čase sa Δ blíži k 0,1 V. Počiatočná hodnota je Δ μ H +> 0,25 V.
V opačnom smere sa nachádza aj H + -ATP-syntetáza, ktorá je v chloroplastoch rozpoznaná ako komplex "CF 1 + F 0". Hlava її (F 1) žasne nad názvom chloroplastu nad strómou. Protón vyshtovuyutsya cez CF 0 + F 1 s matricou zvanou, že v aktívnom centre F 1 tvrdí ATP pre energiu protónového potenciálu.
Na základe formy mitochondriálneho kopijníka v tylaktoide є, mabut, zbavenom dvoch získaných diamantov, syntéza jednej molekuly ATP potrebuje nahradiť dva tri protóny, t.j. produkcia 3 H + / 1 mol AT.
Teraz, v prvej fáze fotosyntézy, pred hodinou svetelných reakcií, sú ATP a NADP vytvorené chloroplastom strómy. · N - produkty, potrebné pre tmavé reakcie.


Mechanizmus temného štádia fotosyntézy |
Tmavé reakcie na fotosyntézu - proces začlenenia oxidu uhličitého do organickej hmoty s prijatím sacharidov (fotosyntéza glukózy z CO 2). Reakcie prebiehajú proti chloroplastom v stróme za účasti produktov vo svetelnom štádiu fotosyntézy - ATP a NADP · H2.
Asimilácia na oxid uhličitý (fotochemická karboxylácia) je cyklický proces, ktorý sa nazýva aj pentózofosfátový fotosyntetický cyklus alebo Calvinov cyklus (obr. 7). Môžete vidieť tri hlavné fázy:
Ribulóza-5-fosfát (tsukor, na pomstenie 5 atómov v uhlíku, s prebytkom fosfátu v uhlíku v polohe 5) sa podáva fosforylovaný pre rakhunok ATP, aby sa vytvoril ribulóza difosfát, kým nebude schválený. Posledným slovom je karboxylácia prenosom 2, mabutu, na medziprodukt so šiestimi uhlíkmi, ktorý sa však nepresne odštiepi od pripojených molekúl vody, čím sa tvoria dve molekuly kyseliny fosfoglycerovej. Pretože kyselina fosfoglycerová je zavedená do enzymatickej reakcie, prítomnosť ATP a NADP je potrebná pre ich zdravie · Z tvrdení fosfoglycerolaldehydu (trikarbón zukor - trióza). V dôsledku kondenzácie dvoch takýchto trióz vzniká molekula hexóz, ktorá môže byť zahrnutá do molekuly škrobu a v takom poradí môže byť zaradená do rezervy.
Aby sa dokončila fáza cyklu v procese fotosyntézy, 1 molekula C0 2 sa odreže a pridajú sa 3 molekuly ATP a 4 atómy H (až 2 molekuly NAD · H). Pri hexózofosfáte sa pentózofosfátový cyklus (obr. 8) regeneruje ribulózofosfát, ktorý môže byť použitý ako molekula v oxide uhličitom.
Taktiež popísané reakcie – karboxylácia, obnova či regenerácia – nie je možné použiť špecifickým spôsobom pre fotosyntetické bunky. Jediným rozdielom medzi nimi je skutočnosť, že pre obnovovaciu reakciu sa vytiahnutím kyseliny fosfoglycerovej premení na fosfoglycerolaldehyd, potrebný NADP. · H, nie NAD · N, jak zavzhdi.
Fixácia CO 2 ribulóza difosfátom katalyzovaná enzýmom ribulóza difosfát karboxylázou: ribulóza difosfát + CO 2 -> 3-fosfoglycerát Dal 3-fosfoglycerát podľa doplnkového NADP · H 2 a ATP na glyceraldehyd-3-fosfát. Reakciu katalyzuje enzým – glyceraldehyd-3-fosfátdehydrogenáza. Glyceraldehyd-3-fosfát sa ľahko izomerizuje z dihydroxyacetónfosfátu. V schválenom fruktóza-bisfosfáte je zločin triózafosfátu (zákerná reakcia, ktorá je katalyzovaná fruktóza-bisfosfát-aldolázou). Časť molekúl fruktóza-bisfosfátu sa po ustálení zúčastňuje súčasne s triózo-fosfátmi na regenerácii ribulózadifosfátu (spomalí cyklus) a časť molekúl je škodlivá pre ukladanie sacharidov vo fotosyntetických bunkách, ako je znázornené na diagramoch.
Pidrachovano, na syntézu jednej molekuly glukózy z CO 2 v Calvinovom cykle je potrebných 12 NADP · Н + Н + a 18 ATP (12 molekúl ATP je vitrizovaných na obnovu 3-fosfoglycerátu a 6 molekúl - na reakcie regenerácie ribulózadifosfátu). Minimálny výkon - 3 ATF: 2 NADP · H2.
Je možné vziať do úvahy ducha princípov, ktoré sú základom fotosyntetických a oxidových fosforylátov, navyše fotofosforylácia je biofosforylačný proces, ktorý nie je fosforylovaným oxidom:

Energia svetla - silou fosforylácie a syntézy organických tekutín (S-Н 2) pri fotosyntéze, navpaki, energia oxidovaných organických tekutín - pri fosforylácii oxidov. Tá istá roslini poskytne život tvorom a ich heterotrofným organizmom:

V uhľohydrátoch je možné etablovať sa počas fotosyntézy, aby v uhlíkatých kastách vyvolali numerické organické frázy kolofónie. Dusíkovo-organická reč je prijímaná fotosyntetickými organizmami obnovou anorganických dusičnanov na atmosférický dusík a serca - obnovou síranov na sulfhydrylové skupiny aminokyselín. Fotosyntéza umožní vytvorenie celkového viskózneho života proteínov, nukleových kyselín, sacharidov, lipidov, kofaktorov a množstva produktov sekundárnej syntézy, cenných liečiv, steroidov, steroidov, steroidov
Fotosyntéza bez chlorofylu
Fotosyntéza bez chlorofylu sa vyskytuje u slanomilných baktérií, čo môže spôsobiť pigment citlivý na fialové svetlo. Pomocou pigmentu sa objavila baktéria rhodopsin, ktorá je podobná zelenej purpure sita – rodopsin a následne sietnica. Bakteriorhodopsín, vstreknutý do membrány slanomilných baktérií, je nastavený na membráne na protónový potenciál, ktorý sa premieňa na ATP. V takejto kategórii je bakteriorhodopsín bezchlorofylovou rekonvertujúcou energiou svetla.
Fotosyntéza a call centrum
Fotosyntéza machu je poháňaná oxidom uhličitým v dôsledku prítomnosti svetla. KKD pre fotosyntézu trocha je viac ako 20 % u pestovaných druhov ruženín; V atmosfére rastie približne 0,03 % (obj.) CO 2 s nárastom do 0,1 %, intenzita fotosyntézy a produktivita ruží, takže rast pestovateľských línií s hydrouhličitanmi je dostatočný. Avšak vm_st 2 v potravinách, 1,0%, dáva veľa energie na fotosyntézu. Pre riziko zbavenia pozemských rosníc sa využívajú 3 % celkového CO 2 zemskej atmosféry, čo sa blíži k 20 miliardám ton.Zásobníky syntetizovaného CO 2 v sacharidoch akumulujú až 4 · 10 18 kJ svetelnej energie. . Zvýšenie ceny za kapacitu elektrárne je 40 miliárd kW. Vedľajší produkt fotosyntézy – kizín – je životne dôležitý pre životne dôležité organizmy a vzdušné mikroorganizmy. Zachovať rast línie znamená zachovať život na Zemi.

Účinnosť fotosyntézy
Efektívnosť fotosyntézy sa dá na prvý pohľad posúdiť prostredníctvom časti dvorného ospalého rádia, ktoré sa spotrebuje na spevokole v hodine spevu, aby sa v organických frázach uložilo do zberu. Produktivitu systému je možné hodnotiť pre množstvo organických suchých prejavov, ktoré je možné odmietnuť z jednej plochy na ryk, a výkon v jednotkách hmotnosti (kg) alebo výroby energie (mJ), získaný na hektár na ryk.
Spôsob, ako odložiť, v takom poradí, z oblasti zberača ospalej energie (listov), dlhodobo fungovať, počet dní na trhu s takýmto šikovným osvetlením, ak je možná fotosyntéza na maximálnu rýchlosť, efektívne. Výsledky hodnoty časti ospalého žiarenia (v %), ktoré má k dispozícii roslín (fotosynteticky aktívne žiarenie, FAR), a znalosť hlavných fotochemických a biologických procesov a termodynamická účinnosť umožňujúca znížiť riziko poranenia
Roslin vikoristovuyut svetlo až do ginu od 400 do 700 nm, to znamená, že časť fotosynteticky aktívnej rádioaktivity klesá na 50% všetkého ospalého svetla. Cena intenzity zemského povrchu je 800-1000 W / m2 pre veľmi ospalý deň (zagalom). Maximálna účinnosť premeny energie počas fotosyntézy bola spriemerovaná, praktické je dosiahnuť 5-6%. Hodnotenie je založené na redukcii CO2 v procese, ako aj na základe fyziologických a fyzických strát. Jeden mól viazaného CO 2 vo forme v sacharide vyprodukuje energiu 0,47 MJ a energia mólu kvanta červeného svetla s dodatočným množstvom 680 nm (najčastejšia energia) pri fotení je 0,176. V takom poradí, minimálny počet mólov kvanta červeného svetla, je potrebné, aby 1 mól CO 2 bol 0,47: 0,176 = 2,7. Časť prenosu niektorých elektrónov z olova pre fixáciu jednej molekuly CO 2 však nie je menšia ako osem kvánt svetla, teoretická účinnosť spojenia je 27: 8 = 33 %. Tsі rosrahunki je rozbité na červené svetlo; je jasné, že pri ostrom svetle bude hodnota nižšia.
Pre najkrajšie poľské mysle je účinnosť fixácie v dewline 3%, môže to byť menej ako v krátkom období rastu, keďže to bude pre celý trh ohromujúce, tu bude medzi 1 a 3%.
V skutočnosti, v strednom natiahnutí horniny, účinnosť fotosyntetického opätovného zavedenia energie v zónach s malým podnebím je 0,5-1,3% a v subtropických plodinách - 0,5-2,5%. Pozrite sa na produkt, ktorý je možné merať pre jedinečnú úroveň intenzity ospalého svetla a vysokú účinnosť fotosyntézy, je ľahké vyhodnotiť z grafov, vznášajúcich sa nad obr. 9.
Význam fotosyntézy
| Články k téme: | |
|
Tvir na tému: ako odhadujete hrozno baby jerry?
29. marca 2015 Create D. London pozná milovníka pleti. Zagalny známky ľudí a ssavtsiv
Mám svoju zvláštnosť. Zviazaný smrad z biosociálneho základu Ľudí. Božská láska k Beatrice V dielach Danteho
Pred špeciálnym trikom s Dante, Vergilіy koristuvavsya. Virshi... | |